

Slo1 is the principal potassium channel of human spermatozoa
- PMID: 24137539
- PMCID: PMC3789364
- DOI: 10.7554/eLife.01009
Slo1 is the principal potassium channel of human spermatozoa
Abstract
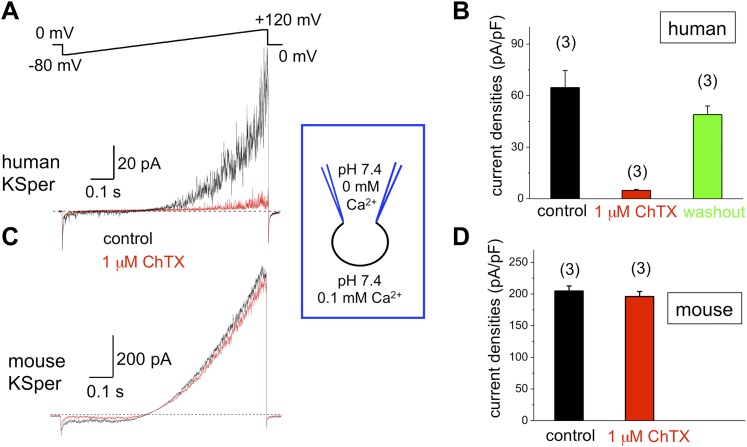
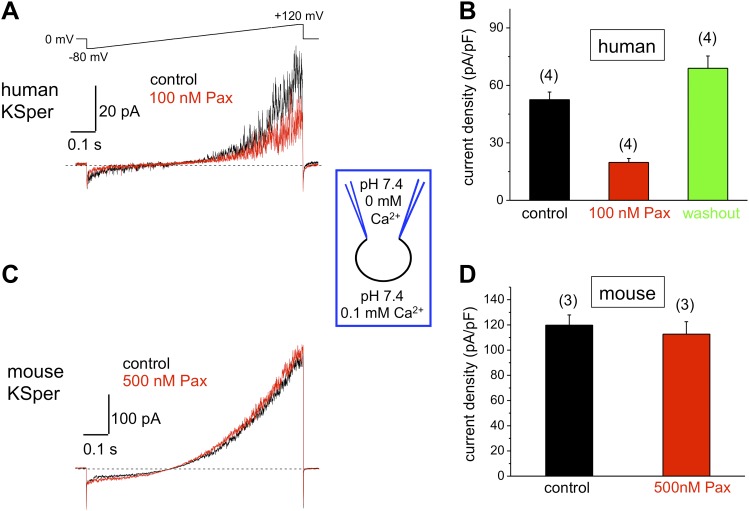
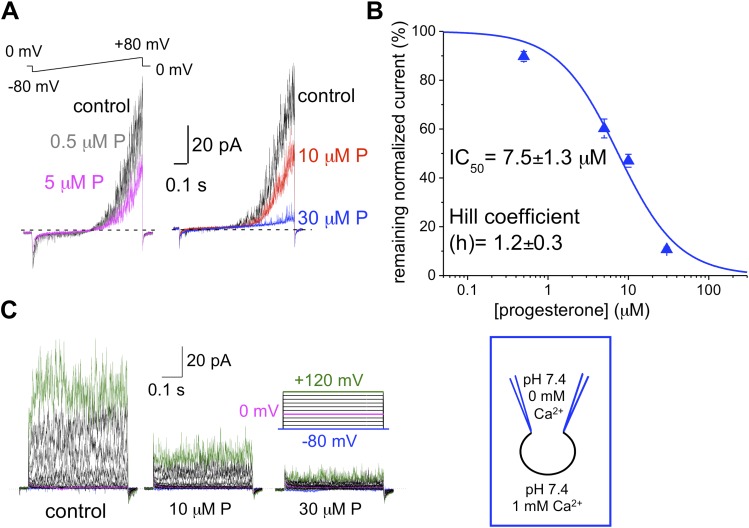
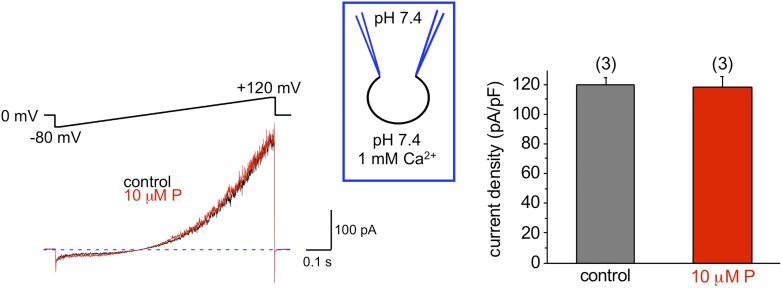
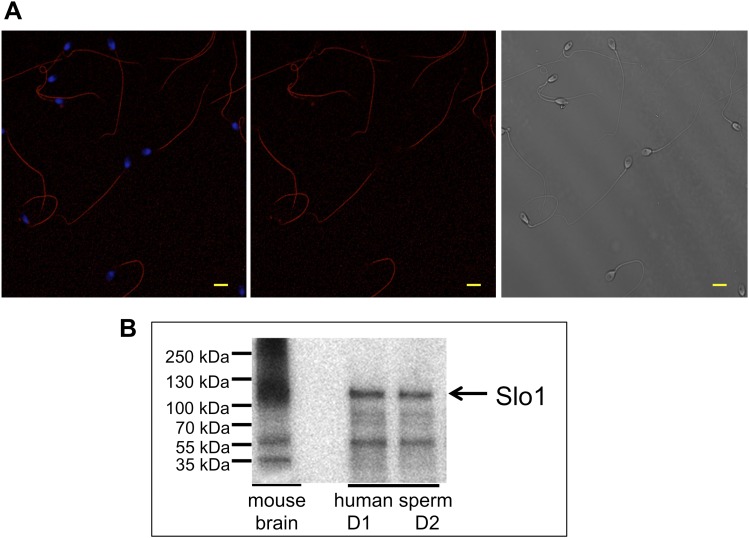
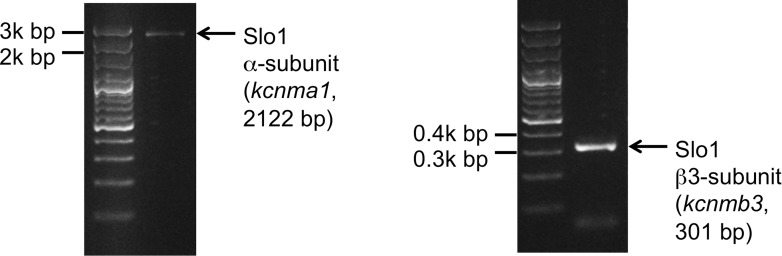
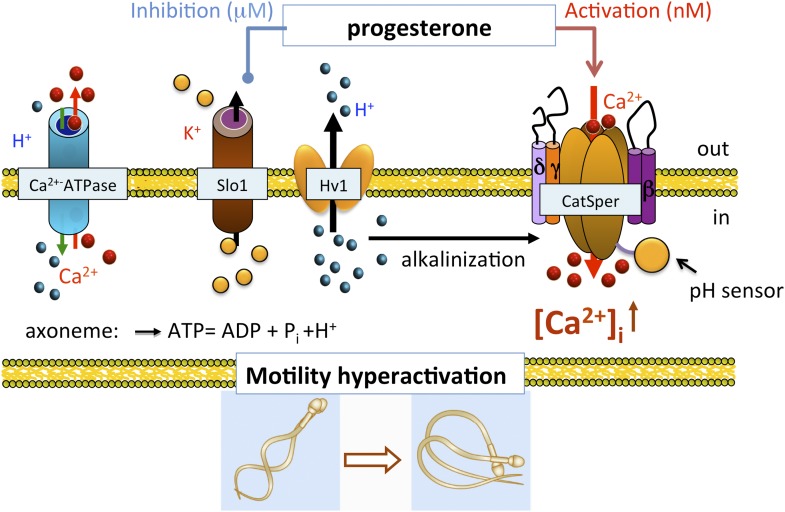
Mammalian spermatozoa gain competence to fertilize an oocyte as they travel through the female reproductive tract. This process is accompanied by an elevation of sperm intracellular calcium and a membrane hyperpolarization. The latter is evoked by K(+) efflux; however, the molecular identity of the potassium channel of human spermatozoa (hKSper) is unknown. Here, we characterize hKSper, reporting that it is regulated by intracellular calcium but is insensitive to intracellular alkalinization. We also show that human KSper is inhibited by charybdotoxin, iberiotoxin, and paxilline, while mouse KSper is insensitive to these compounds. Such unique properties suggest that the Slo1 ion channel is the molecular determinant for hKSper. We show that Slo1 is localized to the sperm flagellum and is inhibited by progesterone. Inhibition of hKSper by progesterone may depolarize the spermatozoon to open the calcium channel CatSper, thus raising [Ca(2+)] to produce hyperactivation and allowing sperm to fertilize an oocyte. DOI:http://dx.doi.org/10.7554/eLife.01009.001.
Keywords: Big Potassium (BK) channel; CatSper; Human; Human KSper; Mouse; Slo1; sperm ion channels; spermatozoa.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
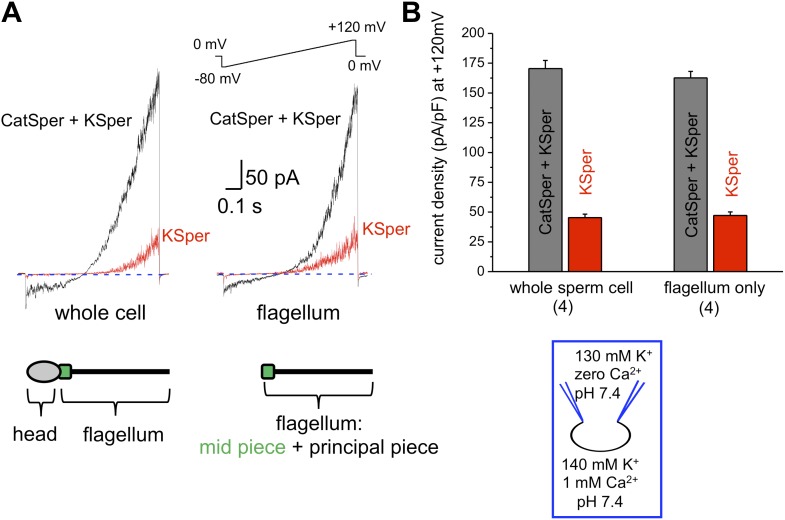
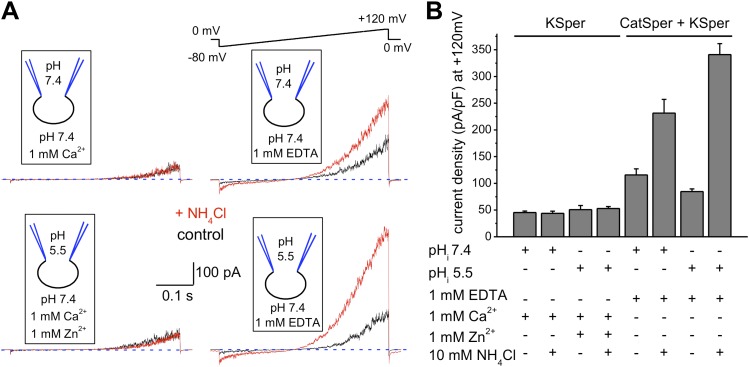
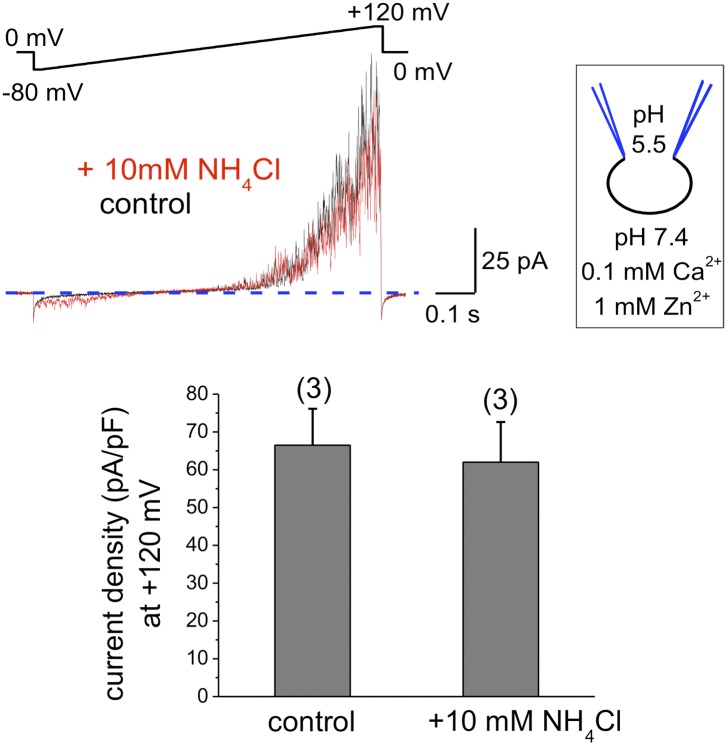
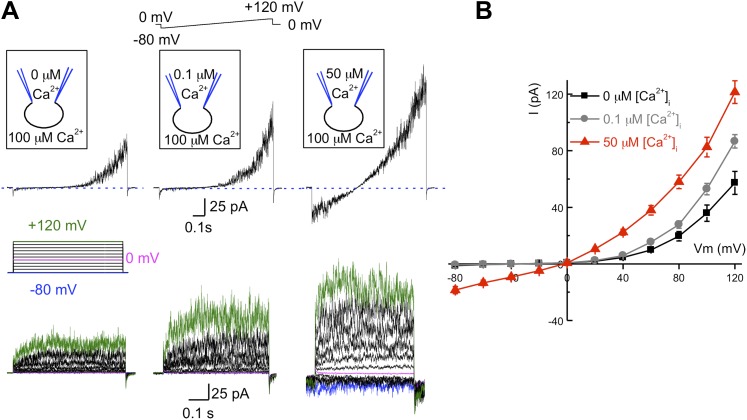

Comment in
-
Sperm BerserKers.Elife. 2013 Oct 8;2:e01469. doi: 10.7554/eLife.01469. Elife. 2013. PMID: 24137547 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
