

Establishment of neurovascular congruency in the mouse whisker system by an independent patterning mechanism
- PMID: 24139045
- PMCID: PMC3998758
- DOI: 10.1016/j.neuron.2013.09.005
Establishment of neurovascular congruency in the mouse whisker system by an independent patterning mechanism
Abstract
Nerves and vessels often run parallel to one another, a phenomenon that reflects their functional interdependency. Previous studies have suggested that neurovascular congruency in planar tissues such as skin is established through a "one-patterns-the-other" model, in which either the nervous system or the vascular system precedes developmentally and then instructs the other system to form using its established architecture as a template. Here, we find that, in tissues with complex three-dimensional structures such as the mouse whisker system, neurovascular congruency does not follow the previous model but rather is established via a mechanism in which nerves and vessels are patterned independently. Given the diversity of neurovascular structures in different tissues, guidance signals emanating from a central organizer in the specific target tissue may act as an important mechanism to establish neurovascular congruency patterns that facilitate unique target tissue function.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
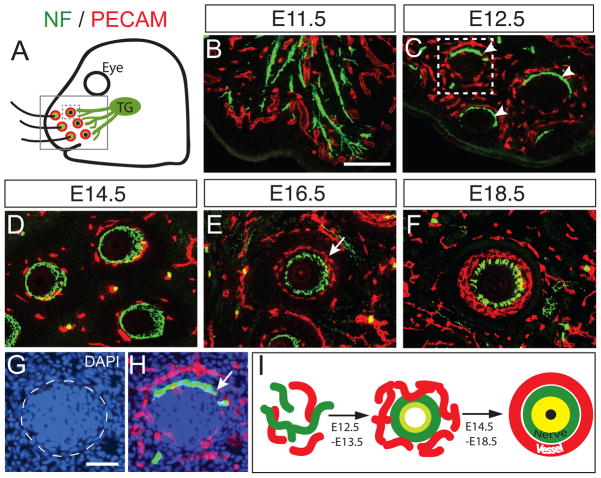
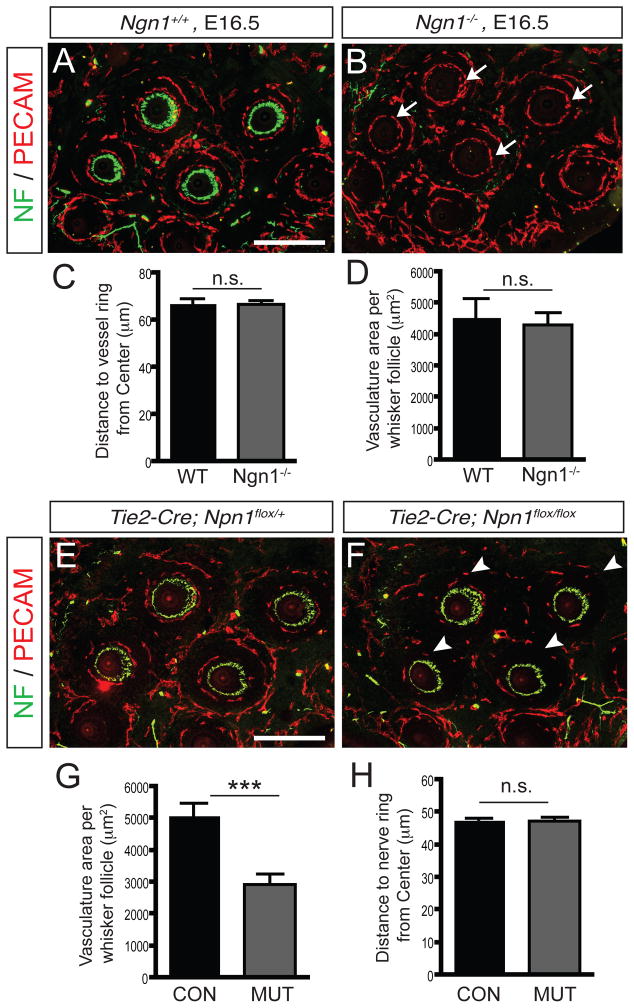
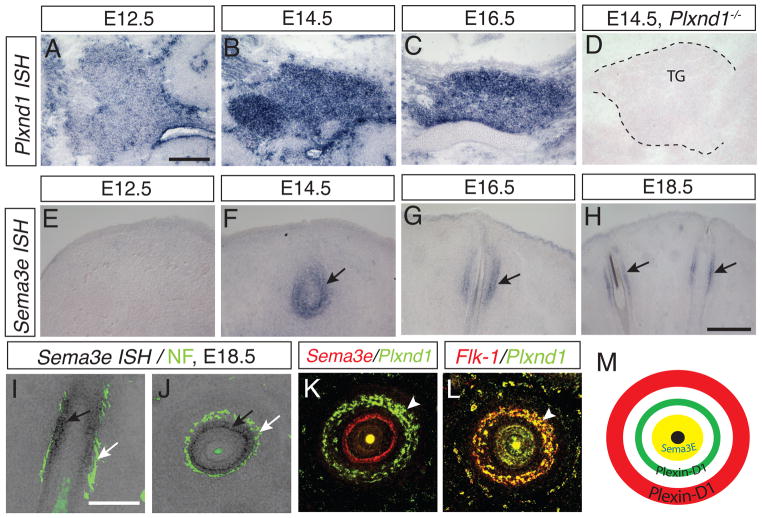
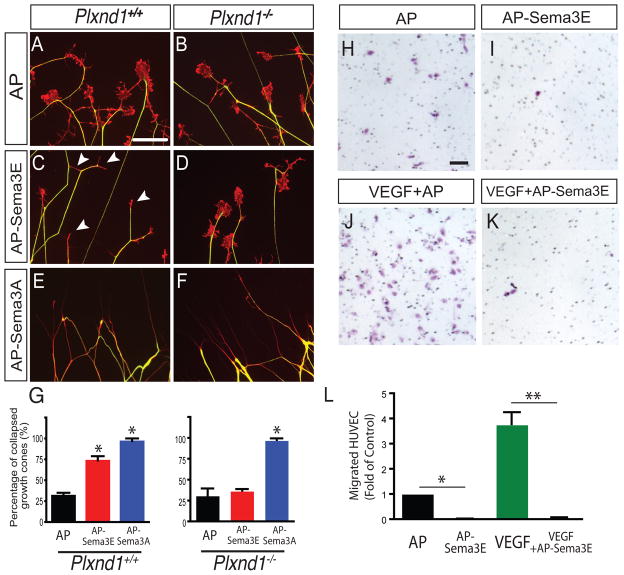
Comment in
-
The declaration of independence of the neurovascular intimacy.Neuron. 2013 Oct 16;80(2):262-5. doi: 10.1016/j.neuron.2013.09.040. Neuron. 2013. PMID: 24139030
Similar articles
-
The declaration of independence of the neurovascular intimacy.Neuron. 2013 Oct 16;80(2):262-5. doi: 10.1016/j.neuron.2013.09.040. Neuron. 2013. PMID: 24139030
-
Neurovascular congruence results from a shared patterning mechanism that utilizes Semaphorin3A and Neuropilin-1.Dev Biol. 2003 Mar 1;255(1):77-98. doi: 10.1016/s0012-1606(02)00045-3. Dev Biol. 2003. PMID: 12618135
-
Specificity of monosynaptic sensory-motor connections imposed by repellent Sema3E-PlexinD1 signaling.Cell Rep. 2013 Nov 14;5(3):748-58. doi: 10.1016/j.celrep.2013.10.005. Epub 2013 Nov 7. Cell Rep. 2013. PMID: 24210822 Free PMC article.
-
Guidance of vascular development: lessons from the nervous system.Circ Res. 2009 Feb 27;104(4):428-41. doi: 10.1161/CIRCRESAHA.108.188144. Circ Res. 2009. PMID: 19246687 Review.
-
Neuronal clues to vascular guidance.Exp Cell Res. 2006 Mar 10;312(5):668-75. doi: 10.1016/j.yexcr.2005.11.009. Epub 2005 Dec 5. Exp Cell Res. 2006. PMID: 16330027 Review.
Cited by
-
Establishment of tooth blood supply and innervation is developmentally regulated and takes place through differential patterning processes.J Anat. 2019 Apr;234(4):465-479. doi: 10.1111/joa.12950. Epub 2019 Feb 21. J Anat. 2019. PMID: 30793310 Free PMC article.
-
An image-based RNAi screen identifies SH3BP1 as a key effector of Semaphorin 3E-PlexinD1 signaling.J Cell Biol. 2014 May 26;205(4):573-90. doi: 10.1083/jcb.201309004. Epub 2014 May 19. J Cell Biol. 2014. PMID: 24841563 Free PMC article.
-
Neuronal and vascular interactions.Annu Rev Neurosci. 2015 Jul 8;38:25-46. doi: 10.1146/annurev-neuro-071714-033835. Epub 2015 Mar 12. Annu Rev Neurosci. 2015. PMID: 25782970 Free PMC article. Review.
-
A Comparative Investigation of Axon-Blood Vessel Growth Interaction in the Regenerating Sciatic and Optic Nerves in Adult Mice.Mol Neurobiol. 2024 Apr;61(4):2215-2227. doi: 10.1007/s12035-023-03705-0. Epub 2023 Oct 21. Mol Neurobiol. 2024. PMID: 37864766
-
PlexinD1 is required for proper patterning of the periocular vascular network and for the establishment of corneal avascularity during avian ocular development.Dev Biol. 2016 Mar 1;411(1):128-39. doi: 10.1016/j.ydbio.2016.01.001. Epub 2016 Jan 16. Dev Biol. 2016. PMID: 26783882 Free PMC article.
References
-
- Armulik A, Genove G, Mae M, Nisancioglu MH, Wallgard E, Niaudet C, He L, Norlin J, Lindblom P, Strittmatter K, et al. Pericytes regulate the blood-brain barrier. Nature. 2010;468:557–561. - PubMed
-
- Bates D, Taylor GI, Minichiello J, Farlie P, Cichowitz A, Watson N, Klagsbrun M, Mamluk R, Newgreen DF. Neurovascular congruence results from a shared patterning mechanism that utilizes Semaphorin3A and Neuropilin-1. Developmental biology. 2003;255:77–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
