

A reciprocal role of prostate cancer on stromal DNA damage
- PMID: 24141771
- PMCID: PMC4121379
- DOI: 10.1038/onc.2013.431
A reciprocal role of prostate cancer on stromal DNA damage
Abstract
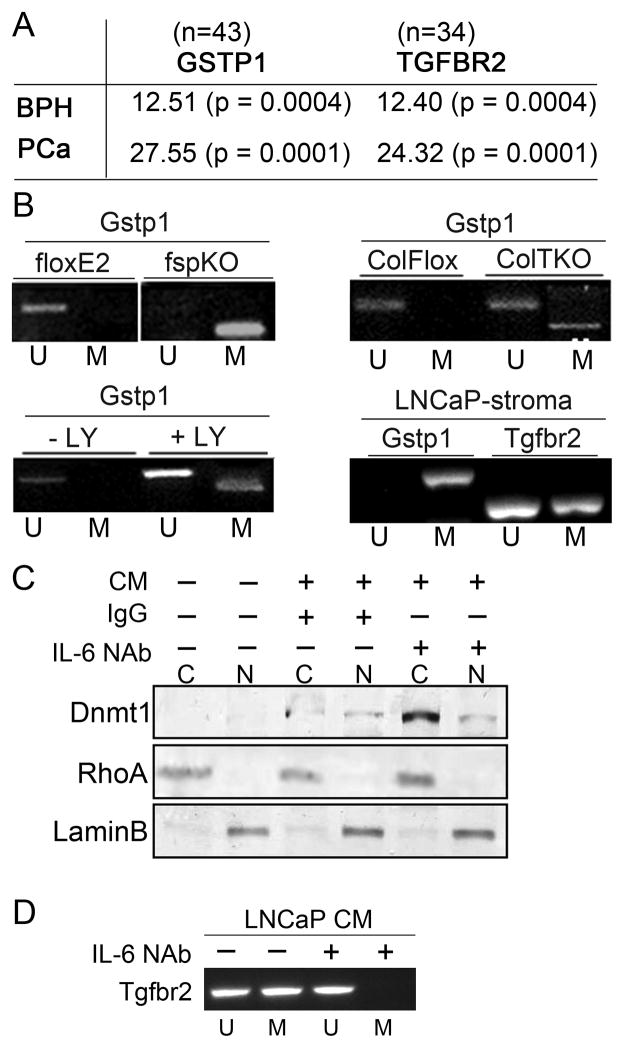
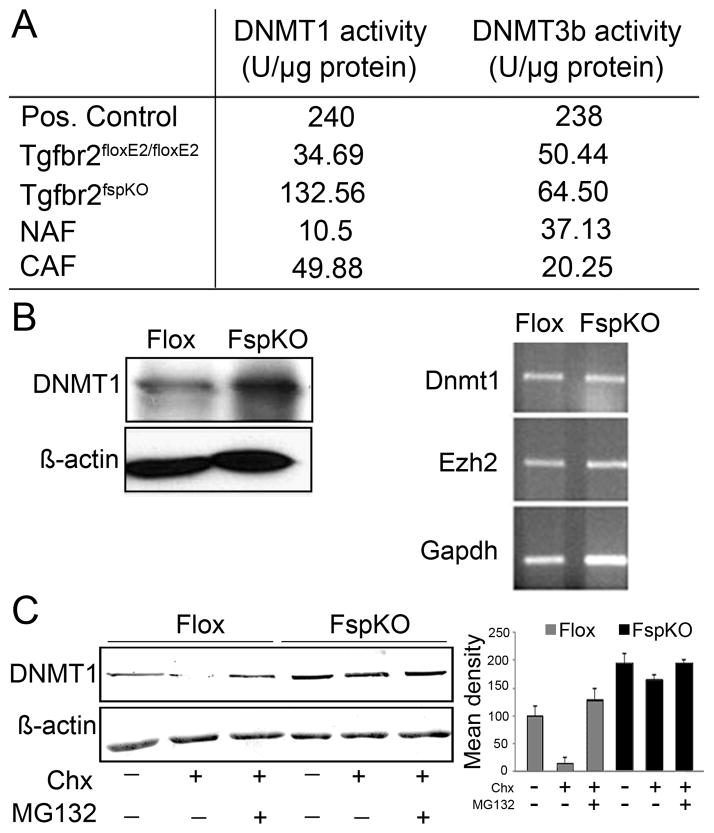
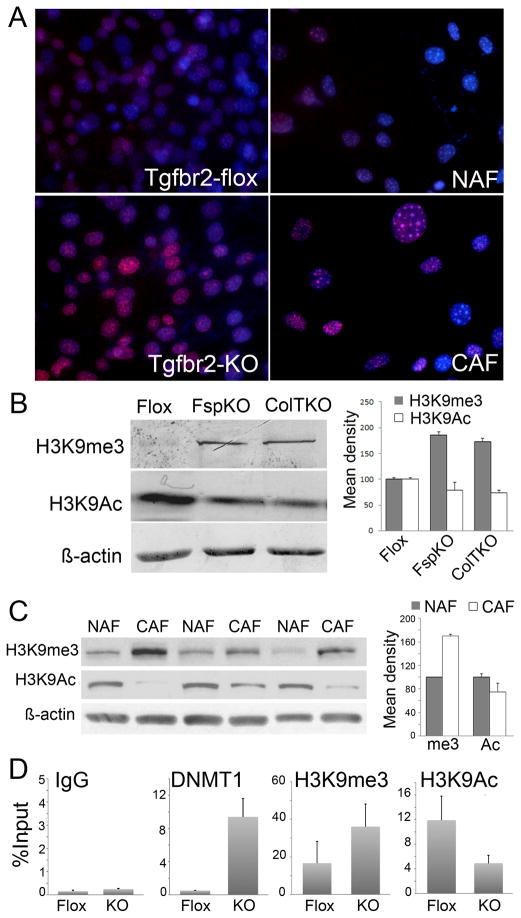
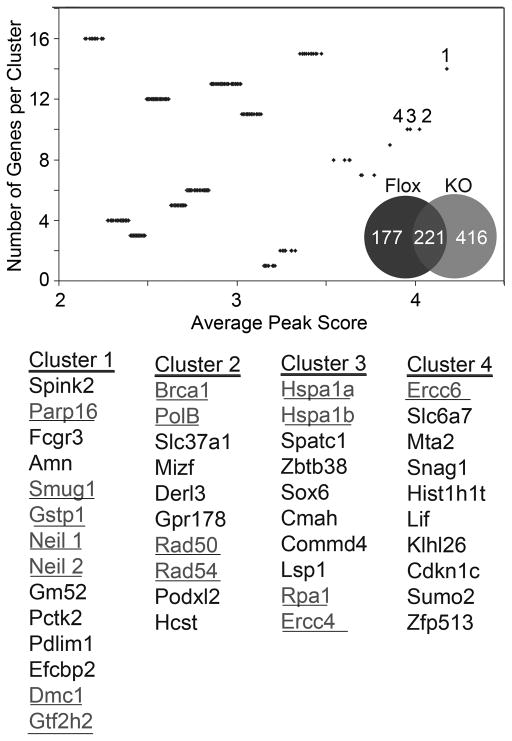
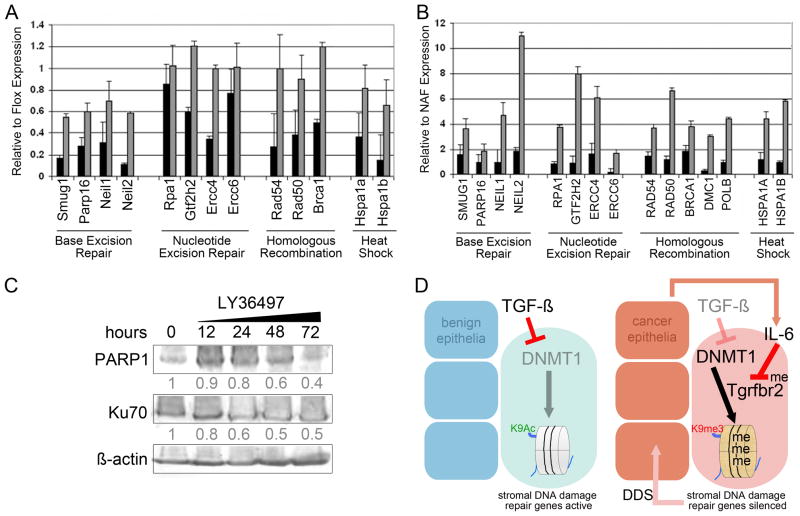
DNA damage found in prostate cancer-associated fibroblasts (CAF) promotes tumor progression. In the absence of somatic mutations in CAF, epigenetic changes dictate how stromal coevolution is mediated in tumors. Seventy percent of prostate cancer patients lose expression of transforming growth factor-beta type II receptor (TGFBR2) in the stromal compartment (n=77, P-value=0.0001), similar to the rate of glutathione S-transferase P1 (GSTP1) silencing. Xenografting of human prostate cancer epithelia, LNCaP, resulted in the epigenetic Tgfbr2 silencing of host mouse prostatic fibroblasts. Stromal Tgfbr2 promoter hypermethylation, initiated by LNCaP cells, was found to be dependent on interleukin 6 expression, based on neutralizing antibody studies. We further found that pharmacologic and transgenic knockout of TGF-β responsiveness in prostatic fibroblasts induced Gstp1 promoter methylation. It is known that TGF-β promotes DNA stability, however, the mechanism is not well understood. Both prostatic human CAF and mouse transgenic knockout of Tgbr2 had elevated DNA methyltransferase I (DNMT1) activity and histone H3 lysine 9 trimethylation (H3K9me3) to suggest greater promoter methylation. Interestingly, the conditional knockout of Tgfbr2 in mouse prostatic fibroblasts, in modeling epigenetic silencing of Tgfbr2, had greater epigenetic gene silencing of multiple DNA damage repair and oxidative stress response genes, based on promoter methylation array analysis. Homologous gene silencing was validated by reverse transcriptase (RT)-PCR in mouse and human prostatic CAF. Not surprisingly, DNA damage repair gene silencing in the prostatic stromal cells corresponded with the presence of DNA damage. Restoring the expression of the epigenetically silenced genes in wild-type fibroblasts with radiation-induced DNA damage reduced tumor progression. Tumor progression was inhibited even when epigenetic silencing was reversed in the Tgfbr2-knockout prostatic fibroblasts. Taken together, fibroblastic epigenetic changes causative of DNA damage, initiated by association with cancer epithelia, is a dominant mediator of tumor progression over TGF-β responsiveness.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Bhowmick NA, Chytil A, Plieth D, Gorska AE, Dumont N, Shappell S, et al. TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science. 2004;303(5659):848–51. Epub 2004/02/07. - PubMed
-
- Nguyen DX, Bos PD, Massague J. Metastasis: from dissemination to organ-specific colonization. Nat Rev Cancer. 2009;9(4):274–84. Epub 2009/03/25. - PubMed
-
- Tuxhorn JA, Ayala GE, Smith MJ, Smith VC, Dang TD, Rowley DR. Reactive stroma in human prostate cancer: induction of myofibroblast phenotype and extracellular matrix remodeling. Clin Cancer Res. 2002;8(9):2912–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
