

An excitable signal integrator couples to an idling cytoskeletal oscillator to drive cell migration
- PMID: 24142103
- PMCID: PMC3838899
- DOI: 10.1038/ncb2859
An excitable signal integrator couples to an idling cytoskeletal oscillator to drive cell migration
Abstract
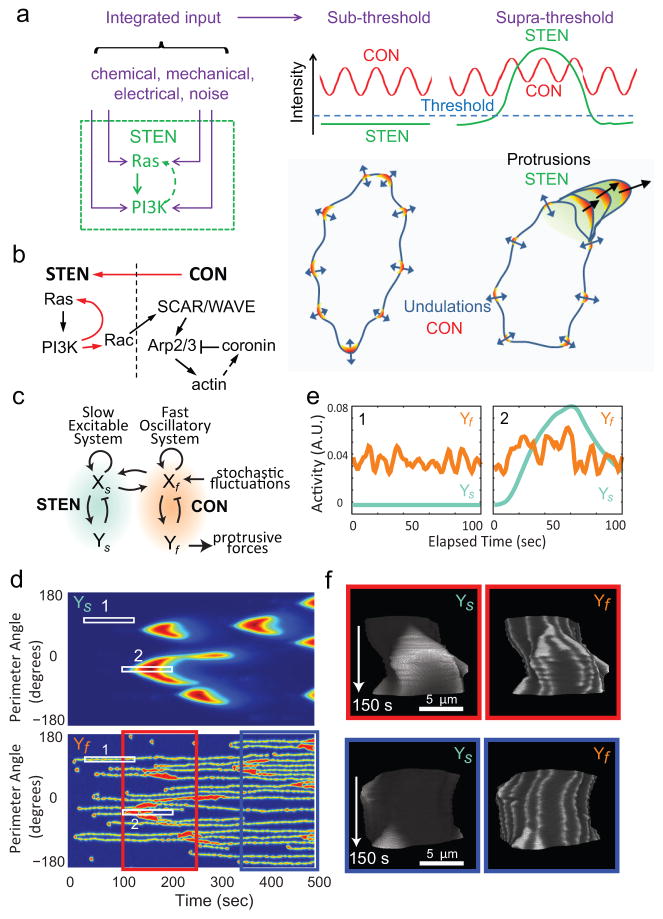
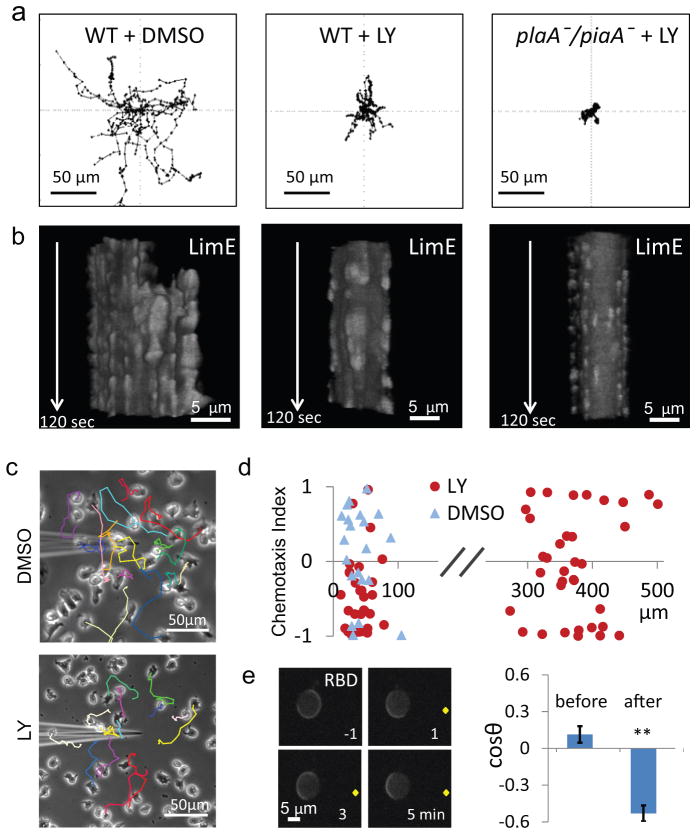
It is generally believed that cytoskeletal activities drive random cell migration, whereas signal transduction events initiated by receptors regulate the cytoskeleton to guide cells. However, we find that the cytoskeletal network, involving SCAR/WAVE, Arp 2/3 and actin-binding proteins, is capable of generating only rapid oscillations and undulations of the cell boundary. The signal transduction network, comprising multiple pathways that include Ras GTPases, PI(3)K and Rac GTPases, is required to generate the sustained protrusions of migrating cells. The signal transduction network is excitable, exhibiting wave propagation, refractoriness and maximal response to suprathreshold stimuli, even in the absence of the cytoskeleton. We suggest that cell motility results from coupling of 'pacemaker' signal transduction and 'idling motor' cytoskeletal networks, and various guidance cues that modulate the threshold for triggering signal transduction events are integrated to control the mode and direction of migration.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
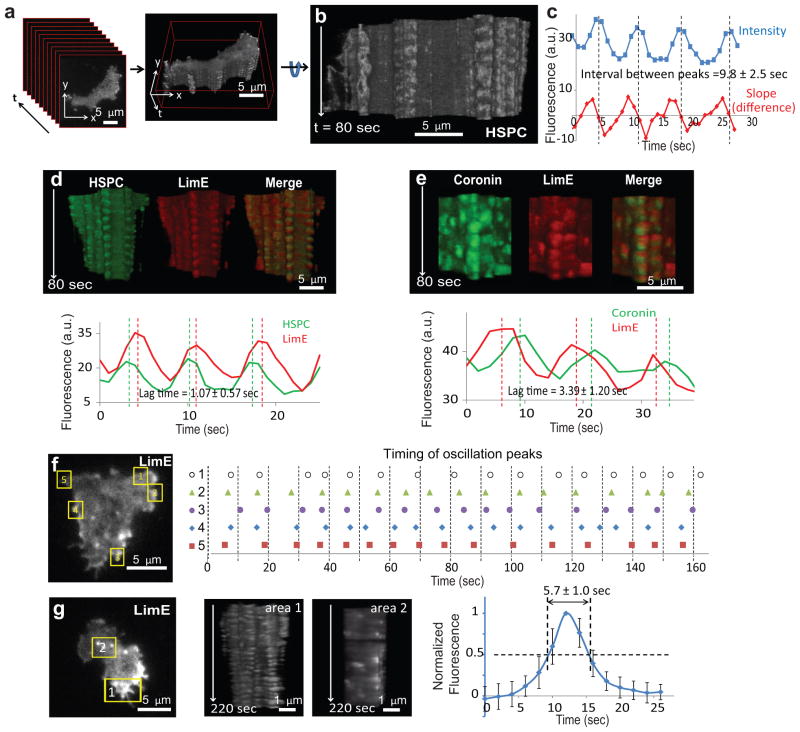
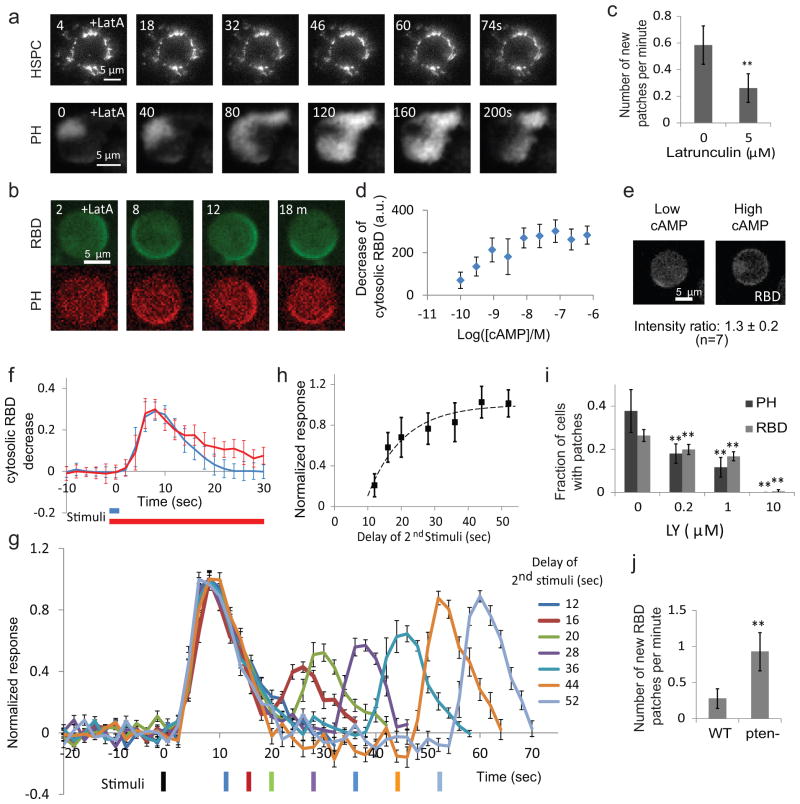
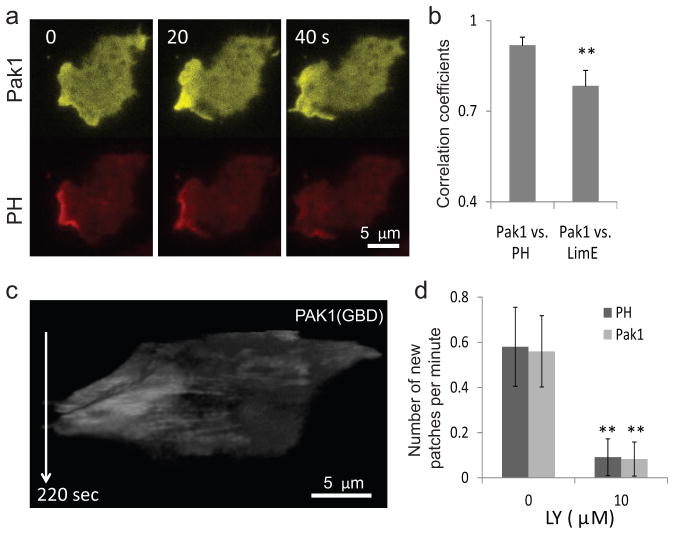
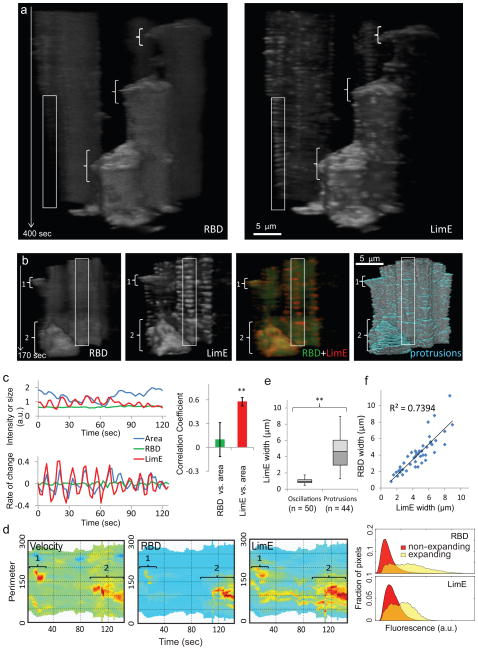
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
