

Role of the twin arginine protein transport pathway in the assembly of the Streptomyces coelicolor cytochrome bc1 complex
- PMID: 24142258
- PMCID: PMC3911139
- DOI: 10.1128/JB.00776-13
Role of the twin arginine protein transport pathway in the assembly of the Streptomyces coelicolor cytochrome bc1 complex
Abstract
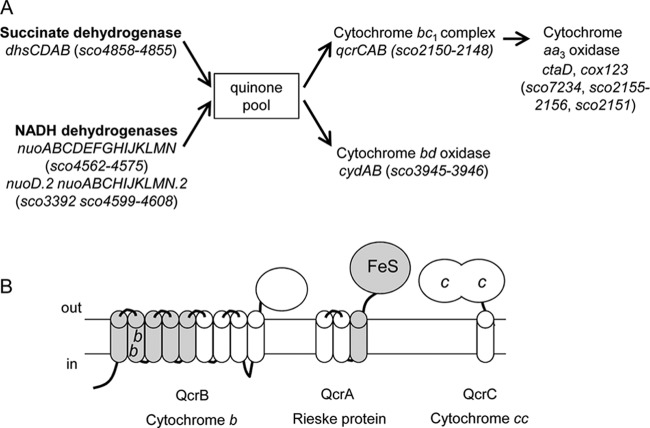
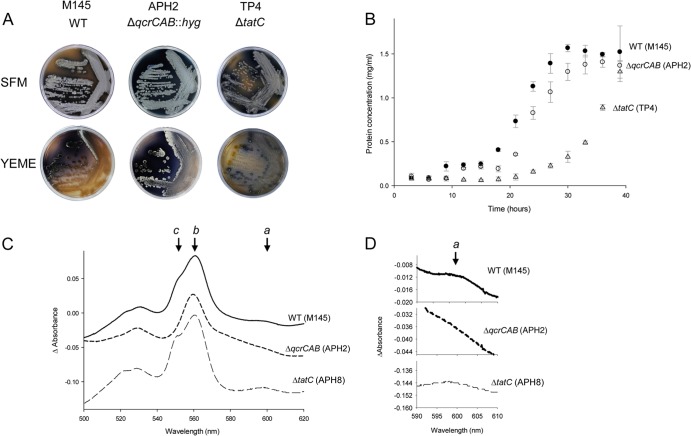
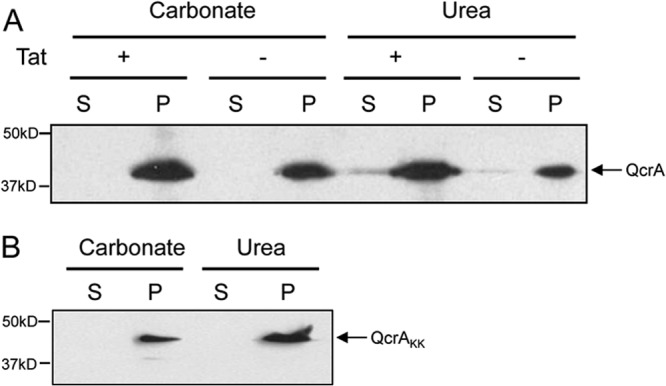
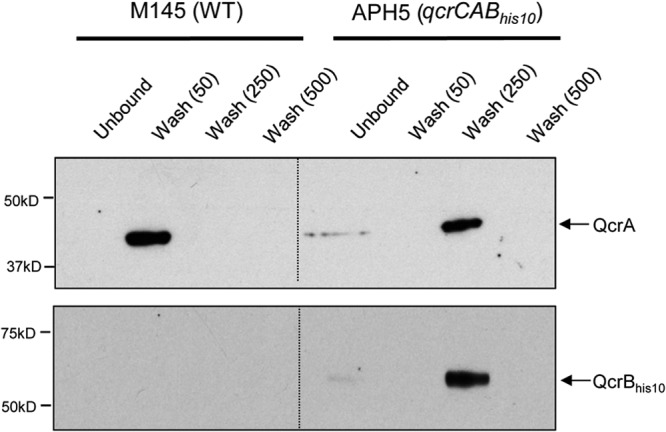
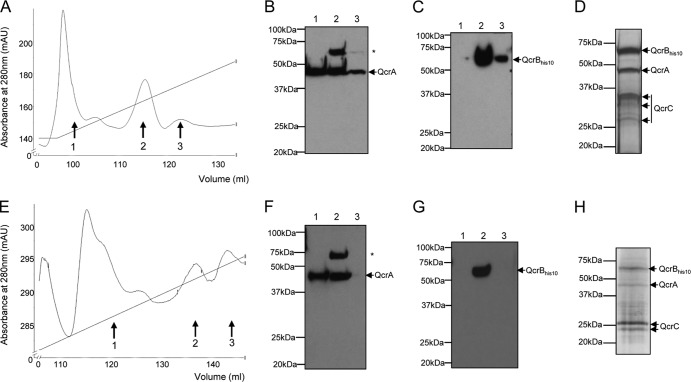
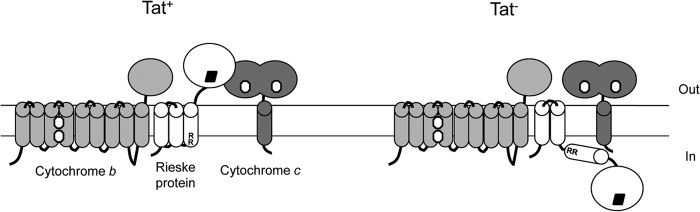
The cytochrome bc1-cytochrome aa3 complexes together comprise one of the major branches of the bacterial aerobic respiratory chain. In actinobacteria, the cytochrome bc1 complex shows a number of unusual features in comparison to other cytochrome bc1 complexes. In particular, the Rieske iron-sulfur protein component of this complex, QcrA, is a polytopic rather than a monotopic membrane protein. Bacterial Rieske proteins are usually integrated into the membrane in a folded conformation by the twin arginine protein transport (Tat) pathway. In this study, we show that the activity of the Streptomyces coelicolor M145 cytochrome bc1 complex is dependent upon an active Tat pathway. However, the polytopic Rieske protein is still integrated into the membrane in a ΔtatC mutant strain, indicating that a second protein translocation machinery also participates in its assembly. Difference spectroscopy indicated that the cytochrome c component of the complex was correctly assembled in the absence of the Tat machinery. We show that the intact cytochrome bc1 complex can be isolated from S. coelicolor M145 membranes by affinity chromatography. Surprisingly, a stable cytochrome bc1 complex containing the Rieske protein can be isolated from membranes even when the Tat system is inactive. These findings strongly suggest that the additional transmembrane segments of the S. coelicolor Rieske protein mediate hydrophobic interactions with one or both of the cytochrome subunits.
Figures
Similar articles
-
Co-factor insertion and disulfide bond requirements for twin-arginine translocase-dependent export of the Bacillus subtilis Rieske protein QcrA.J Biol Chem. 2014 May 9;289(19):13124-31. doi: 10.1074/jbc.M113.529677. Epub 2014 Mar 20. J Biol Chem. 2014. PMID: 24652282 Free PMC article.
-
Co-operation between different targeting pathways during integration of a membrane protein.J Cell Biol. 2012 Oct 15;199(2):303-15. doi: 10.1083/jcb.201204149. Epub 2012 Oct 8. J Cell Biol. 2012. PMID: 23045547 Free PMC article.
-
Assembly of the Rieske iron-sulfur subunit of the cytochrome bc1 complex in the Escherichia coli and Rhodobacter sphaeroides membranes independent of the cytochrome b and c1 subunits.Biochemistry. 1993 Jan 19;32(2):628-36. doi: 10.1021/bi00053a031. Biochemistry. 1993. PMID: 8380704
-
Biogenesis of the yeast cytochrome bc1 complex.Biochim Biophys Acta. 2009 Jan;1793(1):89-96. doi: 10.1016/j.bbamcr.2008.04.011. Epub 2008 May 3. Biochim Biophys Acta. 2009. PMID: 18501197 Review.
-
Design and use of photoactive ruthenium complexes to study electron transfer within cytochrome bc1 and from cytochrome bc1 to cytochrome c.Biochim Biophys Acta. 2013 Nov-Dec;1827(11-12):1309-19. doi: 10.1016/j.bbabio.2012.09.002. Epub 2012 Sep 15. Biochim Biophys Acta. 2013. PMID: 22985600 Free PMC article. Review.
Cited by
-
Ferric Citrate Regulator FecR Is Translocated across the Bacterial Inner Membrane via a Unique Twin-Arginine Transport-Dependent Mechanism.J Bacteriol. 2020 Apr 9;202(9):e00541-19. doi: 10.1128/JB.00541-19. Print 2020 Apr 9. J Bacteriol. 2020. PMID: 32015149 Free PMC article.
-
Co-factor insertion and disulfide bond requirements for twin-arginine translocase-dependent export of the Bacillus subtilis Rieske protein QcrA.J Biol Chem. 2014 May 9;289(19):13124-31. doi: 10.1074/jbc.M113.529677. Epub 2014 Mar 20. J Biol Chem. 2014. PMID: 24652282 Free PMC article.
-
A unifying mechanism for the biogenesis of membrane proteins co-operatively integrated by the Sec and Tat pathways.Elife. 2017 May 17;6:e26577. doi: 10.7554/eLife.26577. Elife. 2017. PMID: 28513434 Free PMC article.
-
Cytochrome bd Oxidase Has an Important Role in Sustaining Growth and Development of Streptomyces coelicolor A3(2) under Oxygen-Limiting Conditions.J Bacteriol. 2018 Jul 25;200(16):e00239-18. doi: 10.1128/JB.00239-18. Print 2018 Aug 15. J Bacteriol. 2018. PMID: 29784883 Free PMC article.
-
Targeting of proteins to the twin-arginine translocation pathway.Mol Microbiol. 2020 May;113(5):861-871. doi: 10.1111/mmi.14461. Epub 2020 Feb 20. Mol Microbiol. 2020. PMID: 31971282 Free PMC article. Review.
References
-
- Shi L, Sohaskey CD, Kana BD, Dawes S, North RJ, Mizrahi V, Gennaro ML. 2005. Changes in energy metabolism of Mycobacterium tuberculosis in mouse lung and under in vitro conditions affecting aerobic respiration. Proc. Natl. Acad. Sci. U. S. A. 102:15629–15634. 10.1073/pnas.0507850102 - DOI - PMC - PubMed
-
- Matsoso LG, Kana BD, Crellin PK, Lea-Smith DJ, Pelosi A, Powell D, Dawes SS, Rubin H, Coppel RL, Mizrahi V. 2005. Function of the cytochrome bc1-aa3 branch of the respiratory network in mycobacteria and network adaptation occurring in response to its disruption. J. Bacteriol. 187:6300–6308. 10.1128/JB.187.18.6300-6308.2005 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
