

Cdk5 phosphorylation of ErbB4 is required for tangential migration of cortical interneurons
- PMID: 24142862
- PMCID: PMC4380000
- DOI: 10.1093/cercor/bht290
Cdk5 phosphorylation of ErbB4 is required for tangential migration of cortical interneurons
Abstract
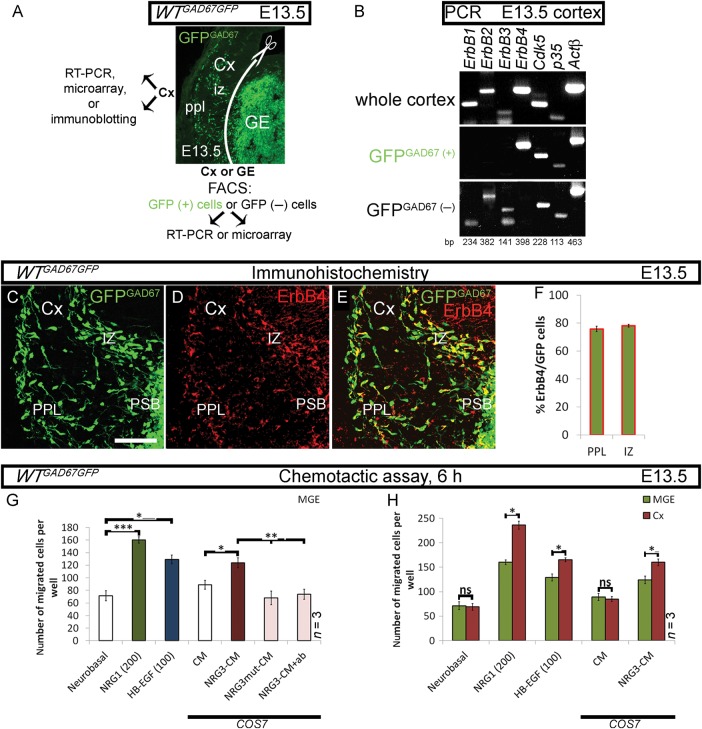
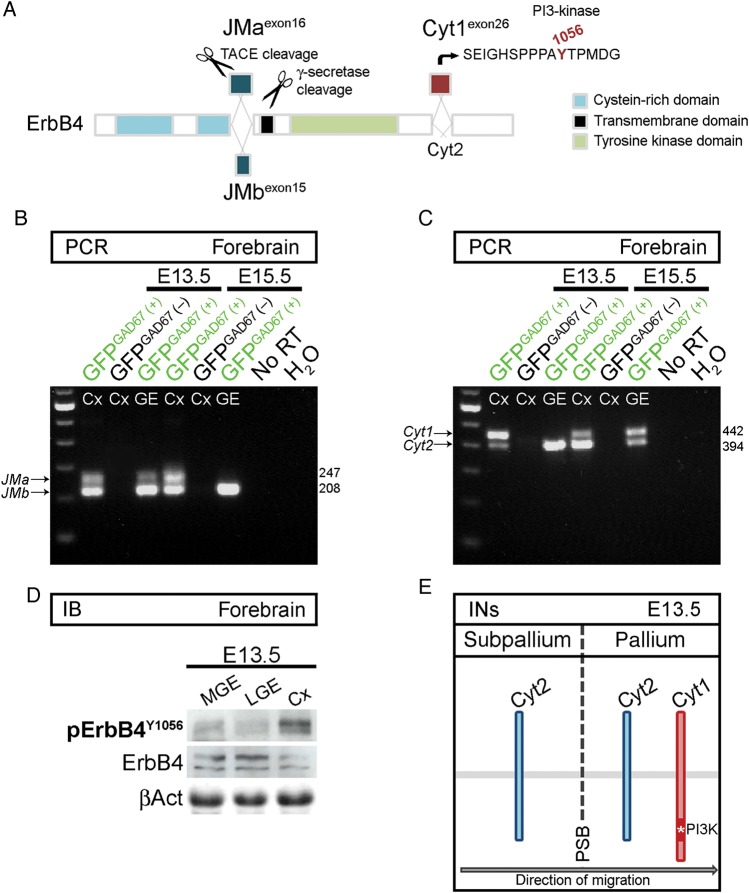
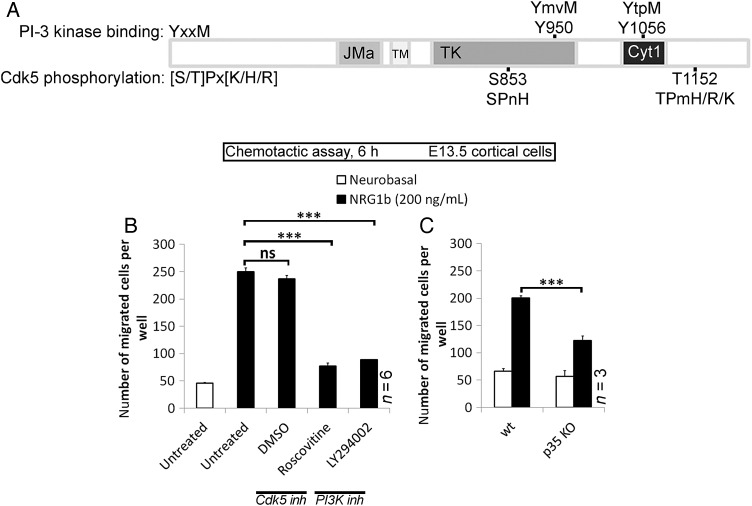
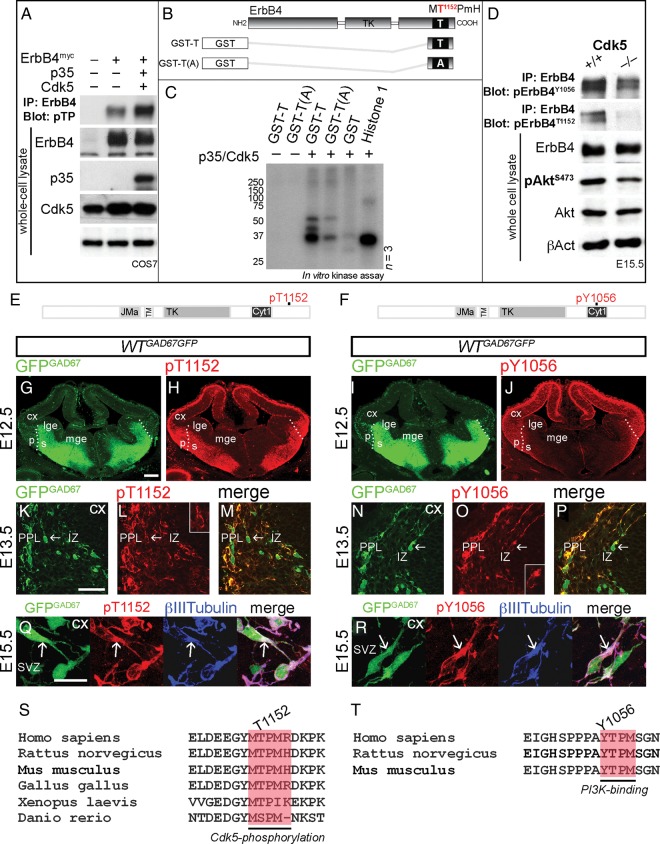
Interneuron dysfunction in humans is often associated with neurological and psychiatric disorders, such as epilepsy, schizophrenia, and autism. Some of these disorders are believed to emerge during brain formation, at the time of interneuron specification, migration, and synapse formation. Here, using a mouse model and a host of histological and molecular biological techniques, we report that the signaling molecule cyclin-dependent kinase 5 (Cdk5), and its activator p35, control the tangential migration of interneurons toward and within the cerebral cortex by modulating the critical neurodevelopmental signaling pathway, ErbB4/phosphatidylinositol 3-kinase, that has been repeatedly linked to schizophrenia. This finding identifies Cdk5 as a crucial signaling factor in cortical interneuron development in mammals.
Keywords: cerebral cortex; interneurons; migration; mouse; phosphorylation.
© The Author 2013. Published by Oxford University Press.
Figures
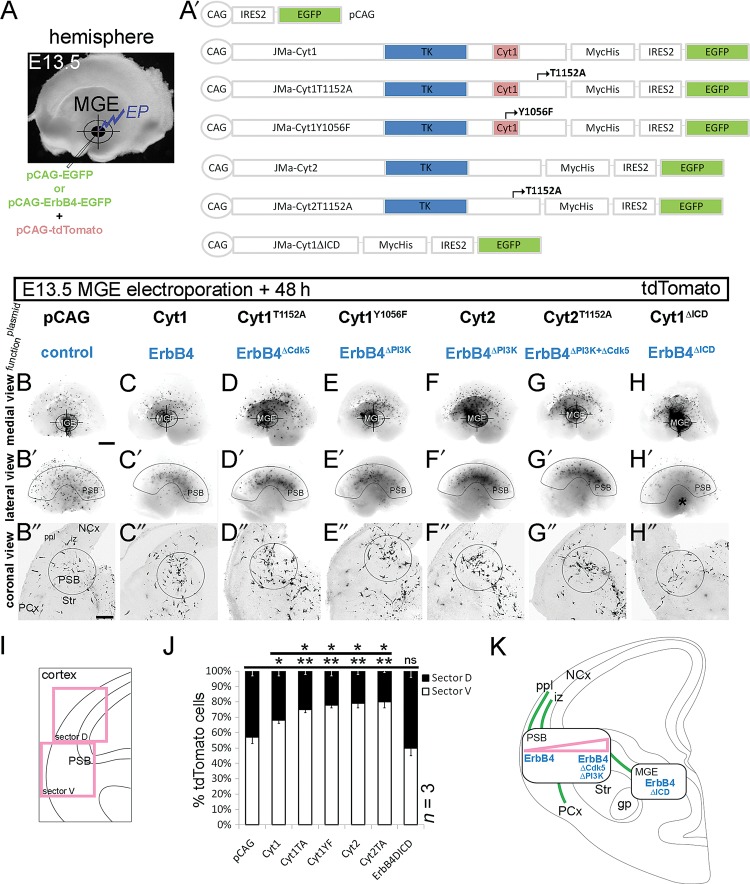
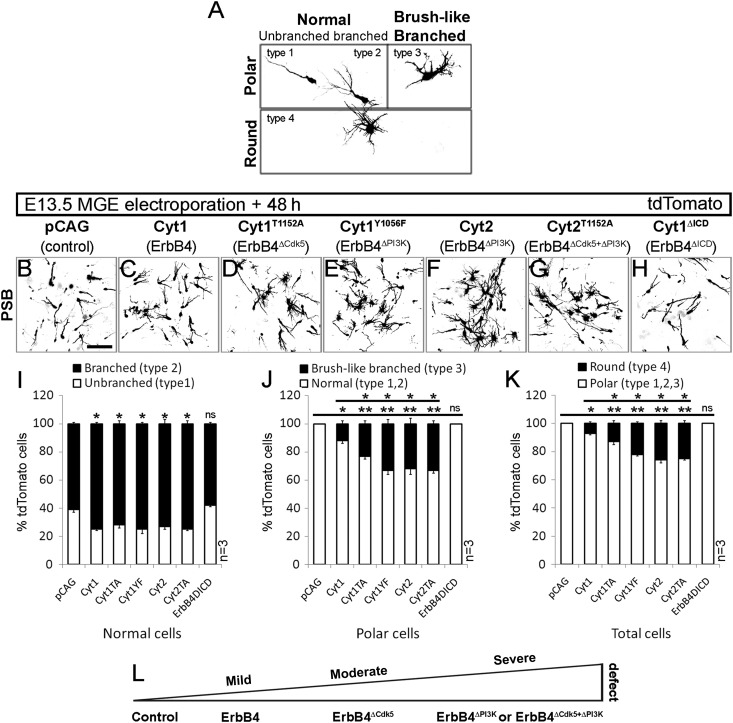
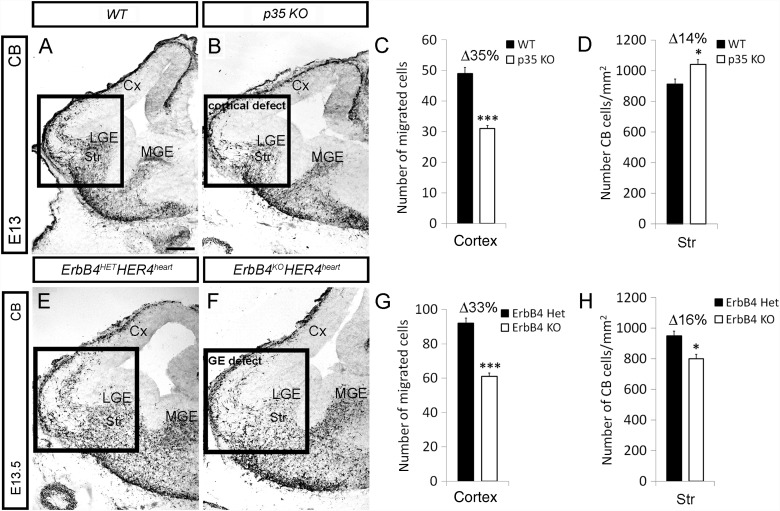
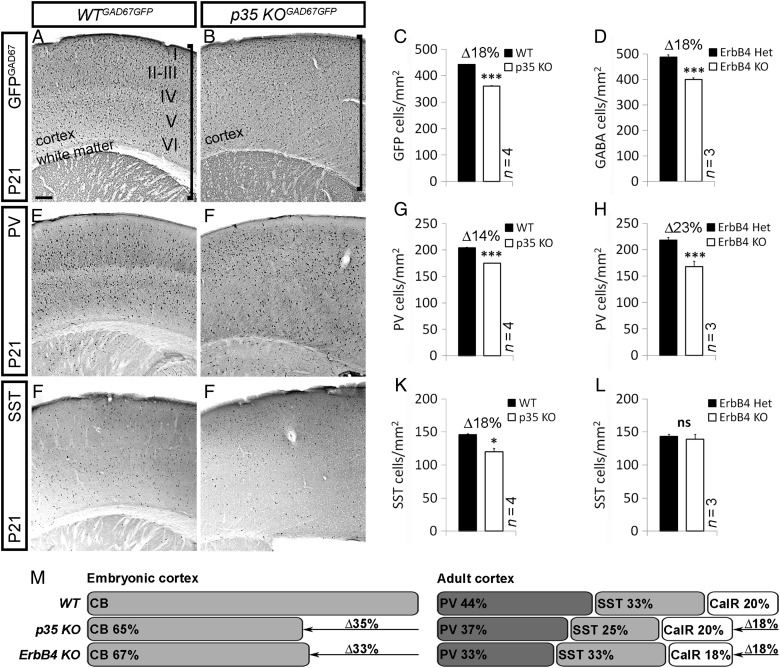
References
-
- Bellion A, Baudoin JP, Alvarez C, Bornens M, Métin C. Nucleokinesis in tangentially migrating neurons comprises two alternating phases: forward migration of the Golgi/centrosome associated with centrosome splitting and myosin contraction at the rear. J Neurosci. 2005;25:5691–5699. doi: 10.1523/JNEUROSCI.1030-05.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
