

Corolla morphology influences diversification rates in bifid toadflaxes (Linaria sect. Versicolores)
- PMID: 24142920
- PMCID: PMC3838546
- DOI: 10.1093/aob/mct214
Corolla morphology influences diversification rates in bifid toadflaxes (Linaria sect. Versicolores)
Abstract
Background and aims: The role of flower specialization in plant speciation and evolution remains controversial. In this study the evolution of flower traits restricting access to pollinators was analysed in the bifid toadflaxes (Linaria sect. Versicolores), a monophyletic group of ~30 species and subspecies with highly specialized corollas.
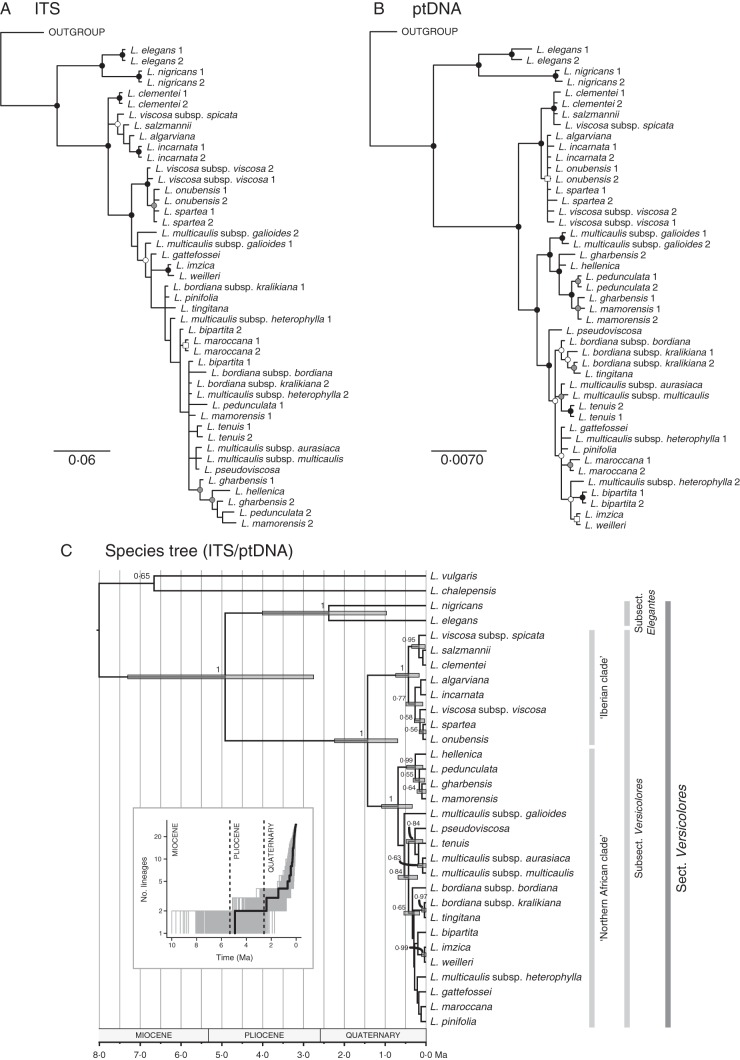
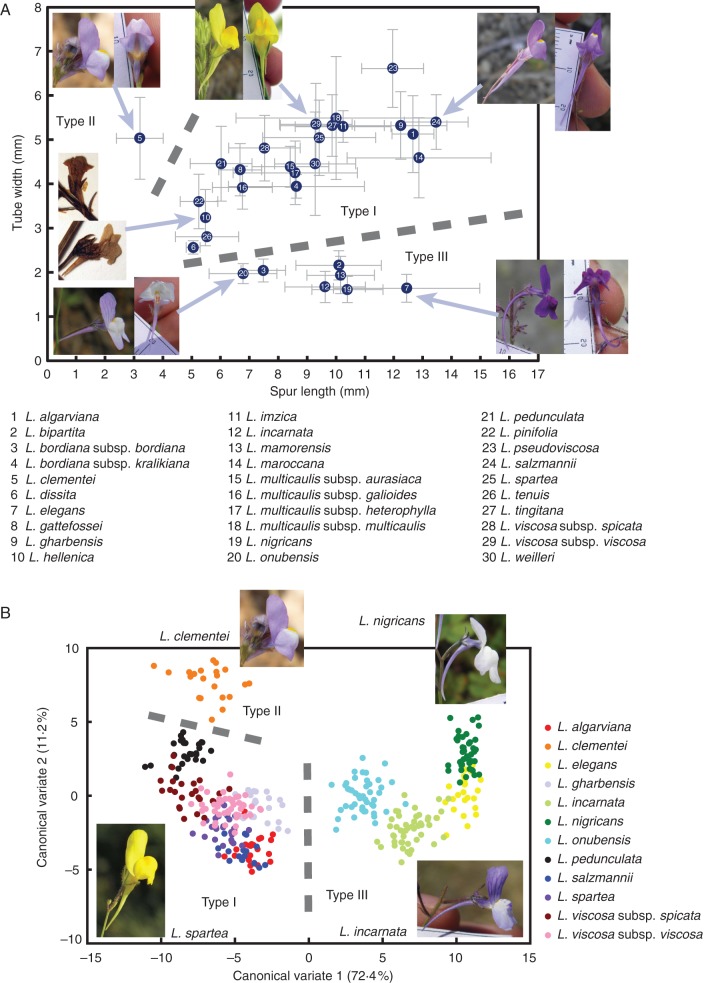
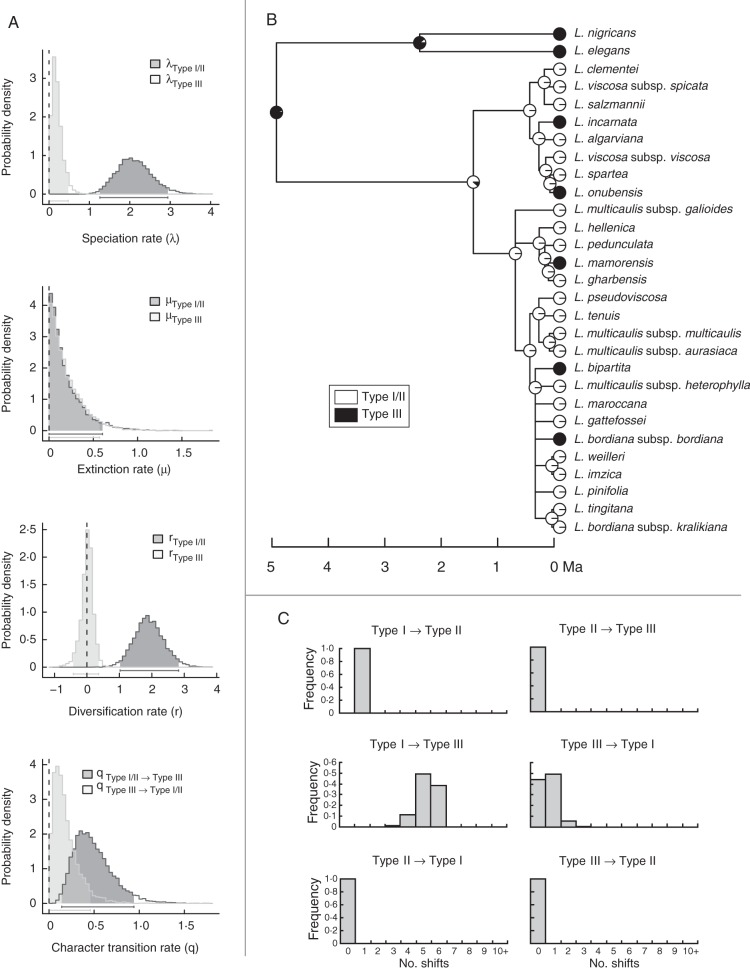
Methods: A time-calibrated phylogeny based on both nuclear and plastid DNA sequences was obtained using a coalescent-based method, and flower morphology was characterized by means of morphometric analyses. Directional trends in flower shape evolution and trait-dependent diversification rates were jointly analysed using recently developed methods, and morphological shifts were reconstructed along the phylogeny. Pollinator surveys were conducted for a representative sample of species.
Key results: A restrictive character state (narrow corolla tube) was reconstructed in the most recent common ancestor of Linaria sect. Versicolores. After its early loss in the most species-rich clade, this character state has been convergently reacquired in multiple lineages of this clade in recent times, yet it seems to have exerted a negative influence on diversification rates. Comparative analyses and pollinator surveys suggest that the narrow- and broad-tubed flowers are evolutionary optima representing divergent strategies of pollen placement on nectar-feeding insects.
Conclusions: The results confirm that different forms of floral specialization can lead to dissimilar evolutionary success in terms of diversification. It is additionally suggested that opposing individual-level and species-level selection pressures may have driven the evolution of pollinator-restrictive traits in bifid toadflaxes.
Keywords: Convergence; Linaria sect. Versicolores; flower specialization; flower tube; nectar spur; pollination; reversal; speciation; species selection; toadflax; trait-dependent diversification.
Figures



Similar articles
-
Bees explain floral variation in a recent radiation of Linaria.J Evol Biol. 2015 Apr;28(4):851-63. doi: 10.1111/jeb.12609. Epub 2015 Mar 16. J Evol Biol. 2015. PMID: 25722058
-
Macroevolutionary dynamics of nectar spurs, a key evolutionary innovation.New Phytol. 2019 Apr;222(2):1123-1138. doi: 10.1111/nph.15654. Epub 2019 Jan 23. New Phytol. 2019. PMID: 30570752
-
Evolution of nectar spur length in a clade of Linaria reflects changes in cell division rather than in cell expansion.Ann Bot. 2018 Nov 3;122(5):801-809. doi: 10.1093/aob/mcx213. Ann Bot. 2018. PMID: 29370374 Free PMC article.
-
Evolvability of flower geometry: Convergence in pollinator-driven morphological evolution of flowers.Semin Cell Dev Biol. 2018 Jul;79:3-15. doi: 10.1016/j.semcdb.2017.09.028. Epub 2017 Oct 7. Semin Cell Dev Biol. 2018. PMID: 28941876 Review.
-
Floral isolation, specialized pollination, and pollinator behavior in orchids.Annu Rev Entomol. 2009;54:425-46. doi: 10.1146/annurev.ento.54.110807.090603. Annu Rev Entomol. 2009. PMID: 19067636 Review.
Cited by
-
Flower color preferences of insects and livestock: effects on Gentiana lutea reproductive success.PeerJ. 2016 Mar 15;4:e1685. doi: 10.7717/peerj.1685. eCollection 2016. PeerJ. 2016. PMID: 27014509 Free PMC article.
-
Climatic and edaphic niche shifts during plant radiation in the Mediterranean biodiversity hotspot.Ann Bot. 2025 Mar 13;135(4):717-734. doi: 10.1093/aob/mcae205. Ann Bot. 2025. PMID: 39673382
-
Adaptation to hummingbird pollination is associated with reduced diversification in Penstemon.Evol Lett. 2019 Aug 2;3(5):521-533. doi: 10.1002/evl3.130. eCollection 2019 Oct. Evol Lett. 2019. PMID: 31636944 Free PMC article.
-
Evolution of habitat preference in 243 species of Bent-toed geckos (Genus Cyrtodactylus Gray, 1827) with a discussion of karst habitat conservation.Ecol Evol. 2020 Nov 22;10(24):13717-13730. doi: 10.1002/ece3.6961. eCollection 2020 Dec. Ecol Evol. 2020. PMID: 33391675 Free PMC article.
-
Contrasting lengths of Pelargonium floral nectar tubes result from late differences in rate and duration of growth.Ann Bot. 2018 Mar 5;121(3):549-560. doi: 10.1093/aob/mcx171. Ann Bot. 2018. PMID: 29293992 Free PMC article.
References
-
- Abdelaziz M, Lorite J, Muñoz-Pajares AJ, Herrador MB, Perfectti F, Gómez JM. Using complementary techniques to distinguish cryptic species: a new Erysimum (Brassicaceae) species from North Africa. American Journal of Botany. 2011;98:1049–1060. - PubMed
-
- Abràmoff MD, Magalhães PJ, Ram SJ. Image processing with ImageJ. Biophotonics International. 2004;11:36–42.
-
- Albrecht GH. Multivariate analysis and the study of form, with special reference to canonical variate analysis. American Zoologist. 1980;20:679–693.
-
- Arendt J, Reznick D. Convergence and parallelism reconsidered: what have we learned about the genetics of adaptation? Trends in Ecology & Evolution. 2008;23:26–32. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
