

Functional coupling between large-conductance potassium channels and Cav3.2 voltage-dependent calcium channels participates in prostate cancer cell growth
- PMID: 24143281
- PMCID: PMC3773341
- DOI: 10.1242/bio.20135215
Functional coupling between large-conductance potassium channels and Cav3.2 voltage-dependent calcium channels participates in prostate cancer cell growth
Abstract
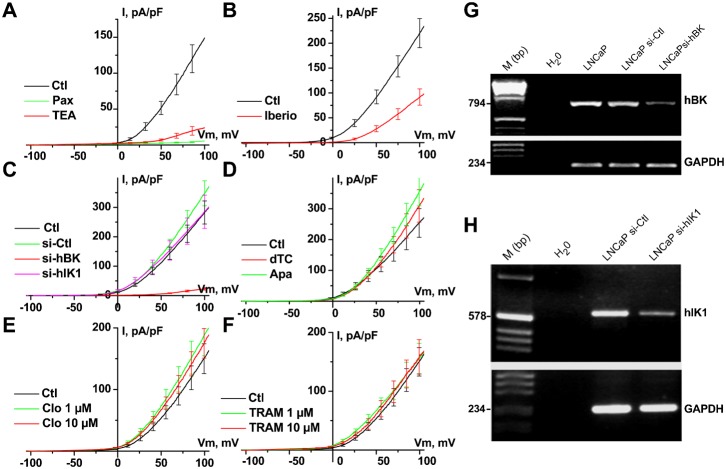
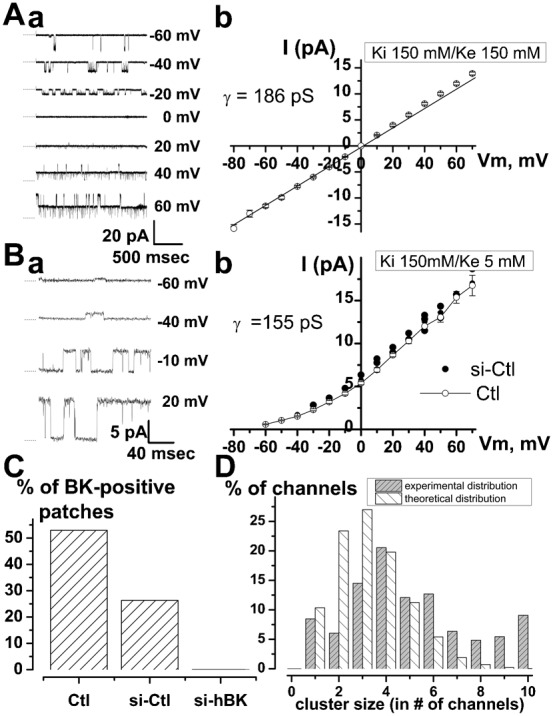
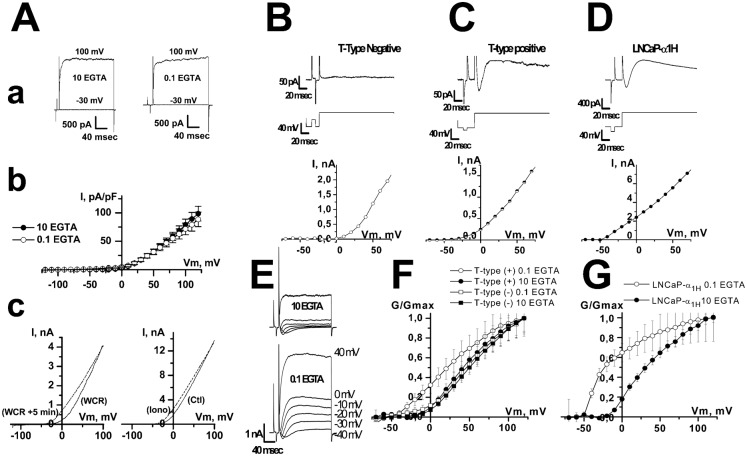
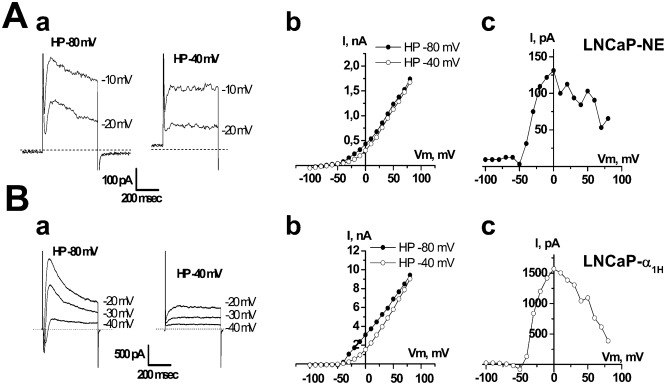
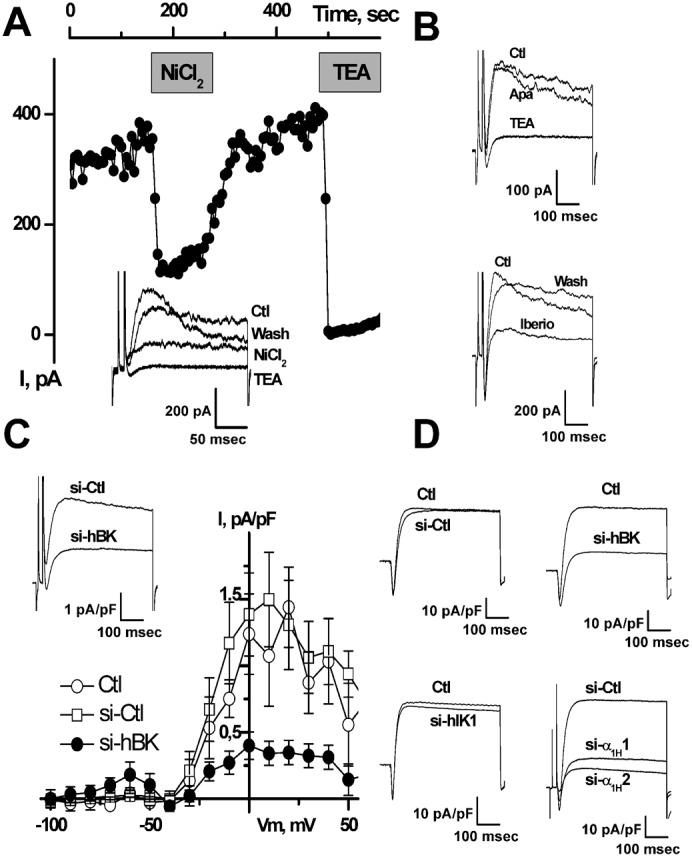
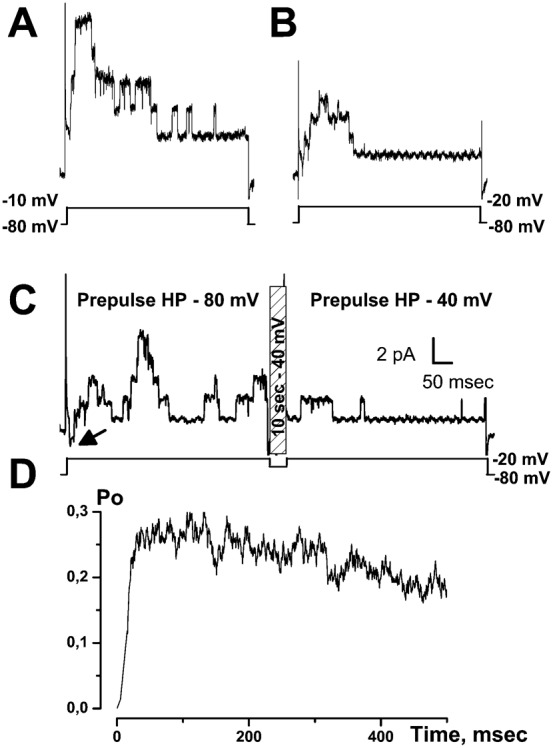
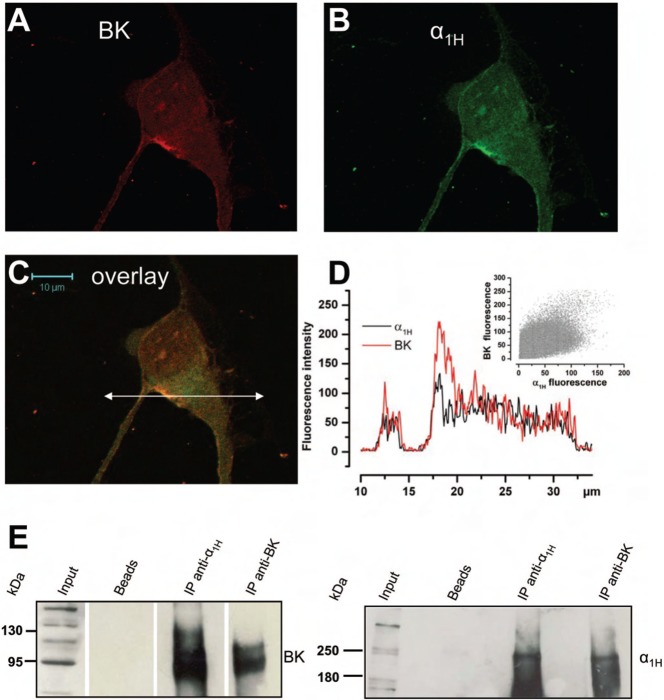
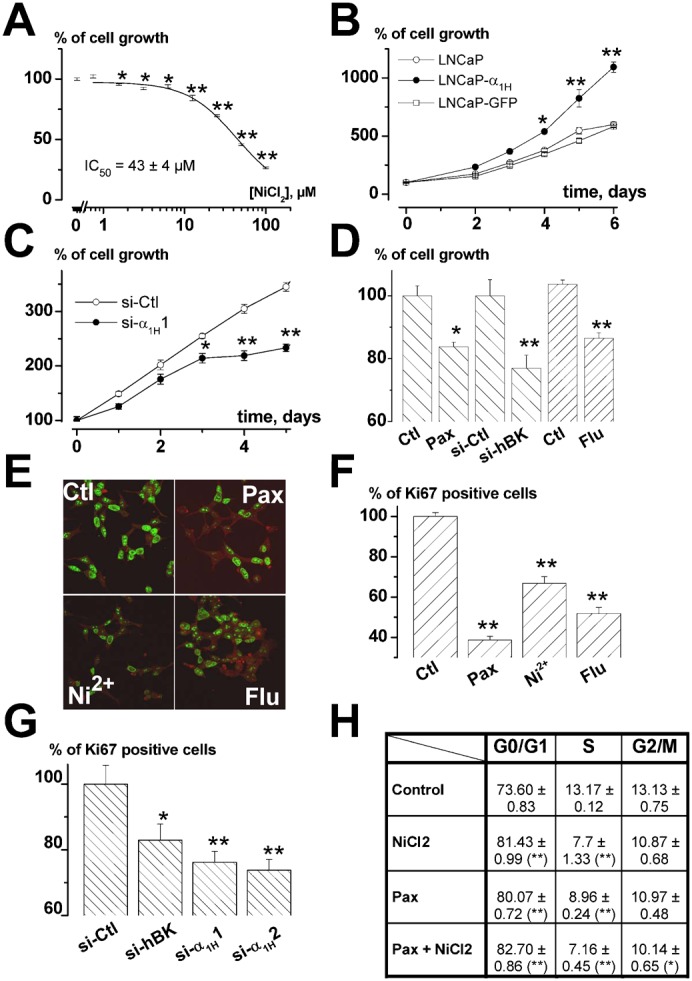
It is strongly suspected that potassium (K(+)) channels are involved in various aspects of prostate cancer development, such as cell growth. However, the molecular nature of those K(+) channels implicated in prostate cancer cell proliferation and the mechanisms through which they control proliferation are still unknown. This study uses pharmacological, biophysical and molecular approaches to show that the main voltage-dependent K(+) current in prostate cancer LNCaP cells is carried by large-conductance BK channels. Indeed, most of the voltage-dependent current was inhibited by inhibitors of BK channels (paxillin and iberiotoxin) and by siRNA targeting BK channels. In addition, we reveal that BK channels constitute the main K(+) channel family involved in setting the resting membrane potential in LNCaP cells at around -40 mV. This consequently promotes a constitutive calcium entry through T-type Cav3.2 calcium channels. We demonstrate, using single-channel recording, confocal imaging and co-immunoprecipitation approaches, that both channels form macromolecular complexes. Finally, using flow cytometry cell cycle measurements, cell survival assays and Ki67 immunofluorescent staining, we show that both BK and Cav3.2 channels participate in the proliferation of prostate cancer cells.
Keywords: BK channels; CACNA1H; Cancer cell growth; Cav3.2; KCa1.1; Proliferation; Prostate; T-type calcium channels.
Conflict of interest statement
Figures
Similar articles
-
Low voltage activation of KCa1.1 current by Cav3-KCa1.1 complexes.PLoS One. 2013 Apr 23;8(4):e61844. doi: 10.1371/journal.pone.0061844. Print 2013. PLoS One. 2013. PMID: 23626738 Free PMC article.
-
Modeling interactions between voltage-gated Ca (2+) channels and KCa1.1 channels.Channels (Austin). 2013 Nov-Dec;7(6):524-9. doi: 10.4161/chan.25867. Epub 2013 Jul 31. Channels (Austin). 2013. PMID: 23928916 Free PMC article.
-
Mechanism of beta4 subunit modulation of BK channels.J Gen Physiol. 2006 Apr;127(4):449-65. doi: 10.1085/jgp.200509436. J Gen Physiol. 2006. PMID: 16567466 Free PMC article.
-
Calcium-activated potassium channels in adrenal chromaffin cells.Ion Channels. 1996;4:261-301. doi: 10.1007/978-1-4899-1775-1_7. Ion Channels. 1996. PMID: 8744211 Review.
-
Large conductance Ca2+-activated K+ (BK) channel: activation by Ca2+ and voltage.Biol Res. 2006;39(3):385-401. doi: 10.4067/s0716-97602006000300003. Epub 2006 Nov 7. Biol Res. 2006. PMID: 17106573 Review.
Cited by
-
Ion Channel Profiling in Prostate Cancer: Toward Cell Population-Specific Screening.Rev Physiol Biochem Pharmacol. 2021;181:39-56. doi: 10.1007/112_2020_22. Rev Physiol Biochem Pharmacol. 2021. PMID: 32737754 Review.
-
CACNA2D2 promotes tumorigenesis by stimulating cell proliferation and angiogenesis.Oncogene. 2015 Oct 16;34(42):5383-94. doi: 10.1038/onc.2014.467. Epub 2015 Jan 26. Oncogene. 2015. PMID: 25619833
-
Ion channels as therapeutic antibody targets.MAbs. 2019 Feb/Mar;11(2):265-296. doi: 10.1080/19420862.2018.1548232. Epub 2018 Dec 10. MAbs. 2019. PMID: 30526315 Free PMC article. Review.
-
Molecular Choreography and Structure of Ca2+ Release-Activated Ca2+ (CRAC) and KCa2+ Channels and Their Relevance in Disease with Special Focus on Cancer.Membranes (Basel). 2020 Dec 15;10(12):425. doi: 10.3390/membranes10120425. Membranes (Basel). 2020. PMID: 33333945 Free PMC article. Review.
-
Evolutionary insights into T-type Ca2+ channel structure, function, and ion selectivity from the Trichoplax adhaerens homologue.J Gen Physiol. 2017 Apr 3;149(4):483-510. doi: 10.1085/jgp.201611683. Epub 2017 Mar 22. J Gen Physiol. 2017. PMID: 28330839 Free PMC article.
References
-
- Bijlenga P., Liu J. H., Espinos E., Haenggeli C. A., Fischer-Lougheed J., Bader C. R., Bernheim L. (2000). T-type alpha 1H Ca2+ channels are involved in Ca2+ signaling during terminal differentiation (fusion) of human myoblasts. Proc. Natl. Acad. Sci. USA 97, 7627–7632 10.1073/pnas.97.13.7627 - DOI - PMC - PubMed
-
- Cambien B., Rezzonico R., Vitale S., Rouzaire-Dubois B., Dubois J. M., Barthel R., Karimdjee B. S., Mograbi B., Schmid-Alliana A., Schmid-Antomarchi H. (2008). Silencing of hSlo potassium channels in human osteosarcoma cells promotes tumorigenesis. Int. J. Cancer 123, 365–371 10.1002/ijc.23511 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
