

Molecular mechanism underlying ethanol activation of G-protein-gated inwardly rectifying potassium channels
- PMID: 24145411
- PMCID: PMC3831446
- DOI: 10.1073/pnas.1311406110
Molecular mechanism underlying ethanol activation of G-protein-gated inwardly rectifying potassium channels
Abstract
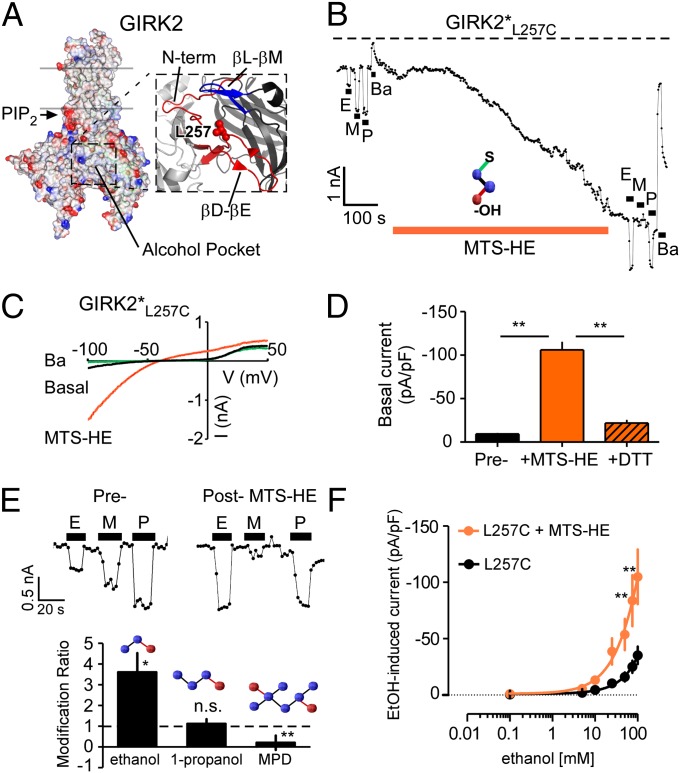
Alcohol (ethanol) produces a wide range of pharmacological effects on the nervous system through its actions on ion channels. The molecular mechanism underlying ethanol modulation of ion channels is poorly understood. Here we used a unique method of alcohol-tagging to demonstrate that alcohol activation of a G-protein-gated inwardly rectifying potassium (GIRK or Kir3) channel is mediated by a defined alcohol pocket through changes in affinity for the membrane phospholipid signaling molecule phosphatidylinositol 4,5-bisphosphate. Surprisingly, hydrophobicity and size, but not the canonical hydroxyl, were important determinants of alcohol-dependent activation. Altering levels of G protein Gβγ subunits, conversely, did not affect alcohol-dependent activation, suggesting a fundamental distinction between receptor and alcohol gating of GIRK channels. The chemical properties of the alcohol pocket revealed here might extend to other alcohol-sensitive proteins, revealing a unique protein microdomain for targeting alcohol-selective therapeutics in the treatment of alcoholism and addiction.
Keywords: Dr-VSP; Kcnj6; Kir3.2; chemical modification; mPhosducin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Hyman SE, Malenka RC, Nestler EJ. Neural mechanisms of addiction: The role of reward-related learning and memory. Annu Rev Neurosci. 2006;29:565–598. - PubMed
-
- Kobayashi T, et al. Ethanol opens G-protein-activated inwardly rectifying K+ channels. Nat Neurosci. 1999;2(12):1091–1097. - PubMed
-
- Lewohl JM, et al. G-protein-coupled inwardly rectifying potassium channels are targets of alcohol action. Nat Neurosci. 1999;2(12):1084–1090. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
