

SIRPα polymorphisms, but not the prion protein, control phagocytosis of apoptotic cells
- PMID: 24145514
- PMCID: PMC3832919
- DOI: 10.1084/jem.20131274
SIRPα polymorphisms, but not the prion protein, control phagocytosis of apoptotic cells
Abstract
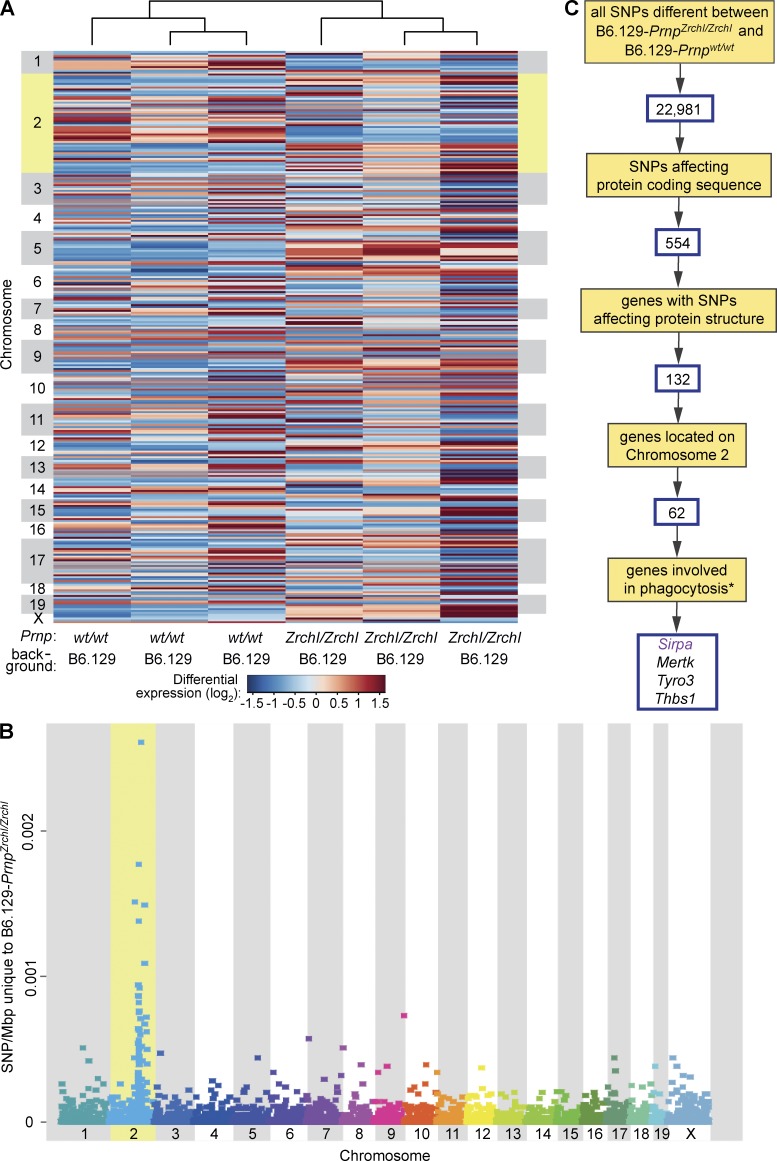
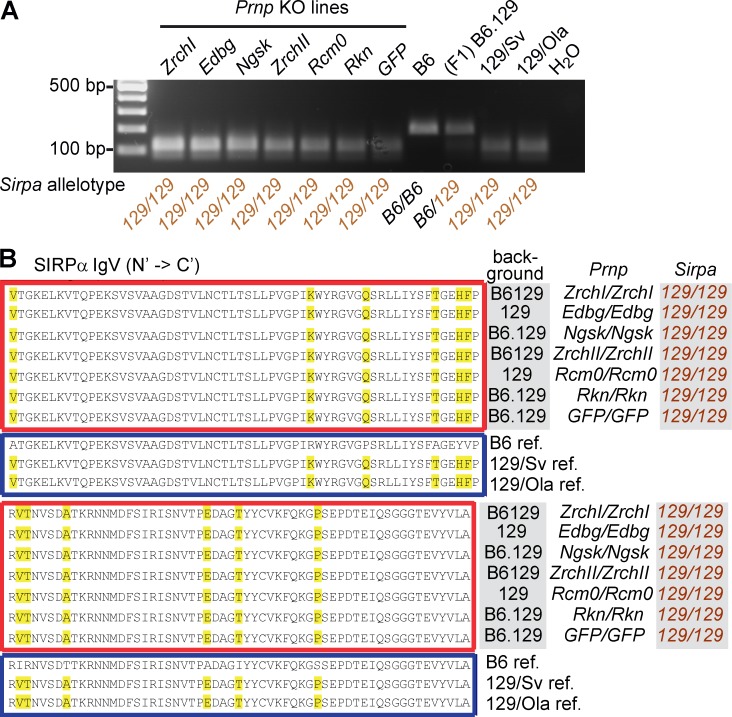
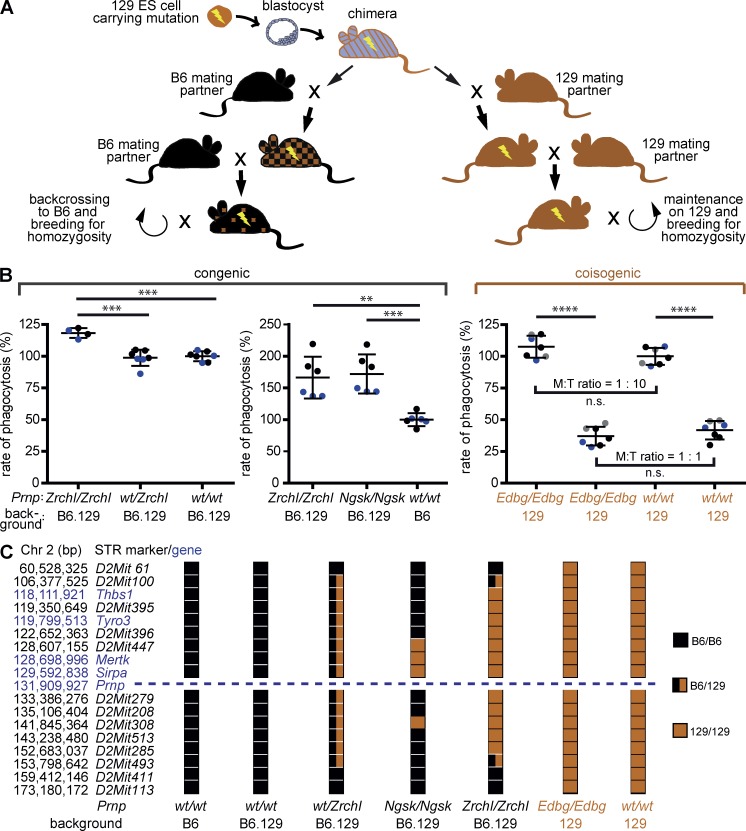
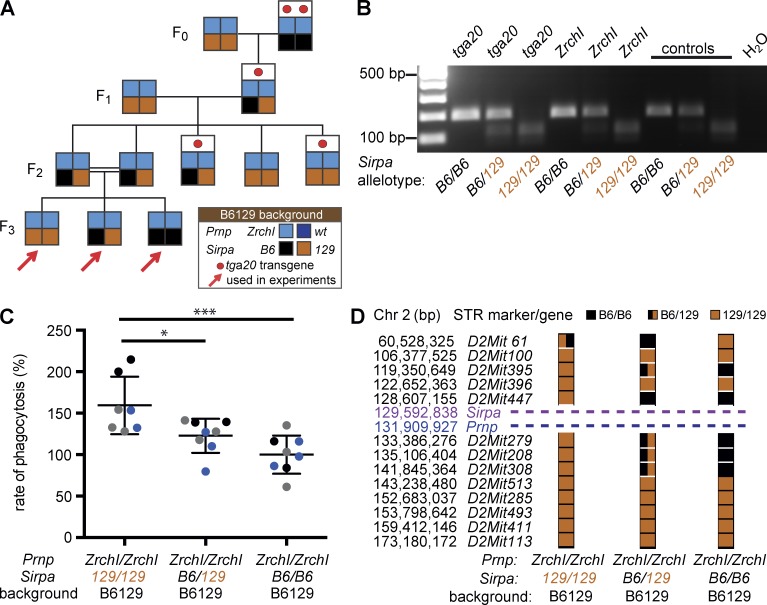
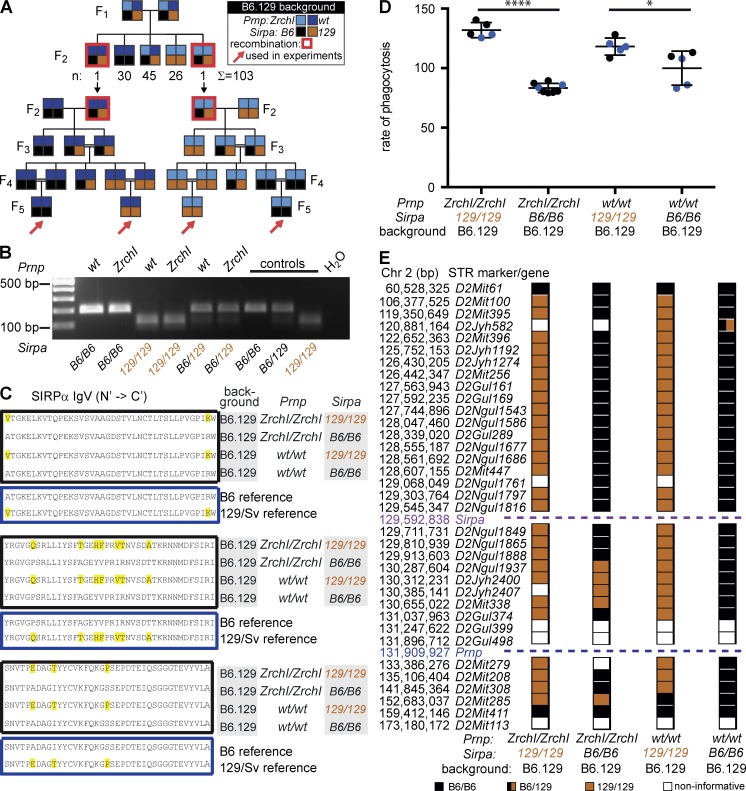
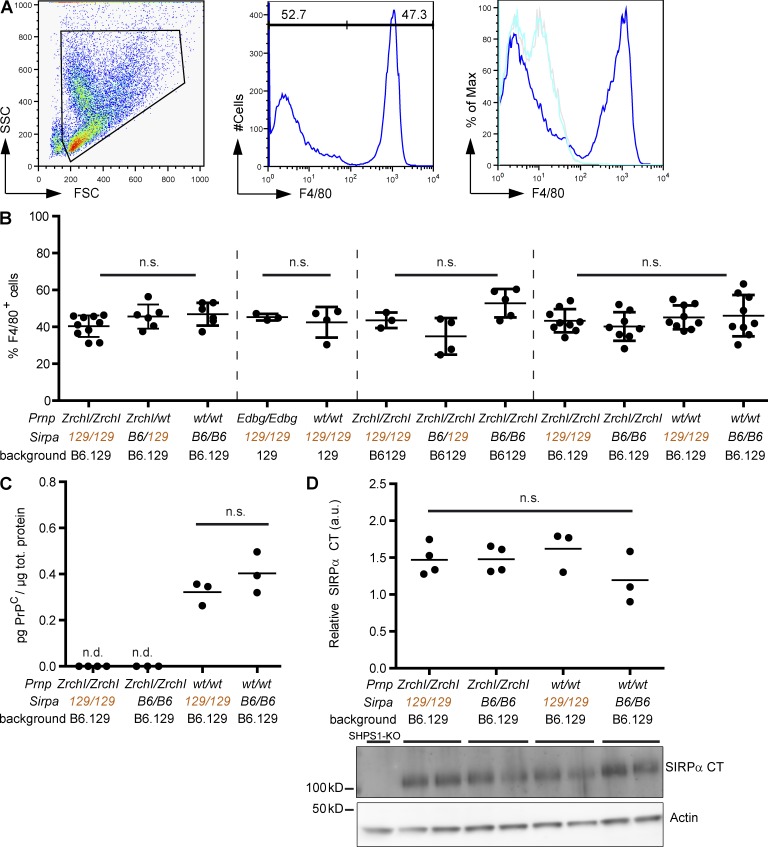
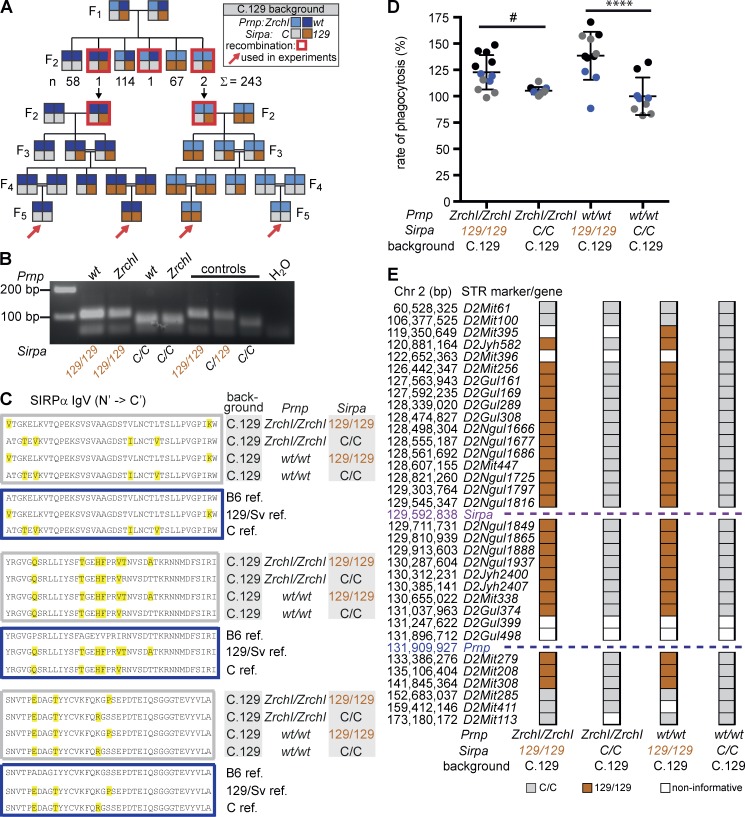
Prnp(-/-) mice lack the prion protein PrP(C) and are resistant to prion infections, but variable phenotypes have been reported in Prnp(-/-) mice and the physiological function of PrP(C) remains poorly understood. Here we examined a cell-autonomous phenotype, inhibition of macrophage phagocytosis of apoptotic cells, previously reported in Prnp(-/-) mice. Using formal genetic, genomic, and immunological analyses, we found that the regulation of phagocytosis previously ascribed to PrP(C) is instead controlled by a linked locus encoding the signal regulatory protein α (Sirpa). These findings indicate that control of phagocytosis was previously misattributed to the prion protein and illustrate the requirement for stringent approaches to eliminate confounding effects of flanking genes in studies modeling human disease in gene-targeted mice. The plethora of seemingly unrelated functions attributed to PrP(C) suggests that additional phenotypes reported in Prnp(-/-) mice may actually relate to Sirpa or other genetic confounders.
Figures
Similar articles
-
Strictly co-isogenic C57BL/6J-Prnp-/- mice: A rigorous resource for prion science.J Exp Med. 2016 Mar 7;213(3):313-27. doi: 10.1084/jem.20151610. Epub 2016 Feb 29. J Exp Med. 2016. PMID: 26926995 Free PMC article.
-
Prion pathogenesis is unaltered in the absence of SIRPα-mediated "don't-eat-me" signaling.PLoS One. 2017 May 17;12(5):e0177876. doi: 10.1371/journal.pone.0177876. eCollection 2017. PLoS One. 2017. PMID: 28545141 Free PMC article.
-
The BALB/c-specific polymorphic SIRPA enhances its affinity for human CD47, inhibiting phagocytosis against human cells to promote xenogeneic engraftment.Exp Hematol. 2014 Mar;42(3):163-171.e1. doi: 10.1016/j.exphem.2013.11.005. Epub 2013 Nov 20. Exp Hematol. 2014. PMID: 24269920
-
The prion gene complex encoding PrP(C) and Doppel: insights from mutational analysis.Gene. 2001 Sep 5;275(1):1-18. doi: 10.1016/s0378-1119(01)00627-8. Gene. 2001. PMID: 11574147 Review.
-
Implications of prion polymorphisms.Prion. 2013 Jul-Aug;7(4):276-9. doi: 10.4161/pri.25566. Epub 2013 Jun 27. Prion. 2013. PMID: 23807178 Free PMC article. Review.
Cited by
-
The role of the NADPH oxidase NOX2 in prion pathogenesis.PLoS Pathog. 2014 Dec 11;10(12):e1004531. doi: 10.1371/journal.ppat.1004531. eCollection 2014 Dec. PLoS Pathog. 2014. PMID: 25502554 Free PMC article.
-
Strictly co-isogenic C57BL/6J-Prnp-/- mice: A rigorous resource for prion science.J Exp Med. 2016 Mar 7;213(3):313-27. doi: 10.1084/jem.20151610. Epub 2016 Feb 29. J Exp Med. 2016. PMID: 26926995 Free PMC article.
-
Loss of prion protein induces a primed state of type I interferon-responsive genes.PLoS One. 2017 Jun 26;12(6):e0179881. doi: 10.1371/journal.pone.0179881. eCollection 2017. PLoS One. 2017. PMID: 28651013 Free PMC article.
-
Ca2+ Signaling but Not Store-Operated Ca2+ Entry Is Required for the Function of Macrophages and Dendritic Cells.J Immunol. 2015 Aug 1;195(3):1202-17. doi: 10.4049/jimmunol.1403013. Epub 2015 Jun 24. J Immunol. 2015. PMID: 26109647 Free PMC article.
-
Frontline Science: Coincidental null mutation of Csf2rα in a colony of PI3Kγ-/- mice causes alveolar macrophage deficiency and fatal respiratory viral infection.J Leukoc Biol. 2017 Feb;101(2):367-376. doi: 10.1189/jlb.4HI0316-157R. Epub 2016 Jul 28. J Leukoc Biol. 2017. PMID: 27468760 Free PMC article.
References
-
- Balducci C., Beeg M., Stravalaci M., Bastone A., Sclip A., Biasini E., Tapella L., Colombo L., Manzoni C., Borsello T., et al. 2010. Synthetic amyloid-beta oligomers impair long-term memory independently of cellular prion protein. Proc. Natl. Acad. Sci. USA. 107:2295–2300 10.1073/pnas.0911829107 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
