

Innate immune signaling induces interleukin-7 production from salivary gland cells and accelerates the development of primary Sjögren's syndrome in a mouse model
- PMID: 24147035
- PMCID: PMC3798297
- DOI: 10.1371/journal.pone.0077605
Innate immune signaling induces interleukin-7 production from salivary gland cells and accelerates the development of primary Sjögren's syndrome in a mouse model
Erratum in
- PLoS One. 2014;9(9):e108573
Abstract
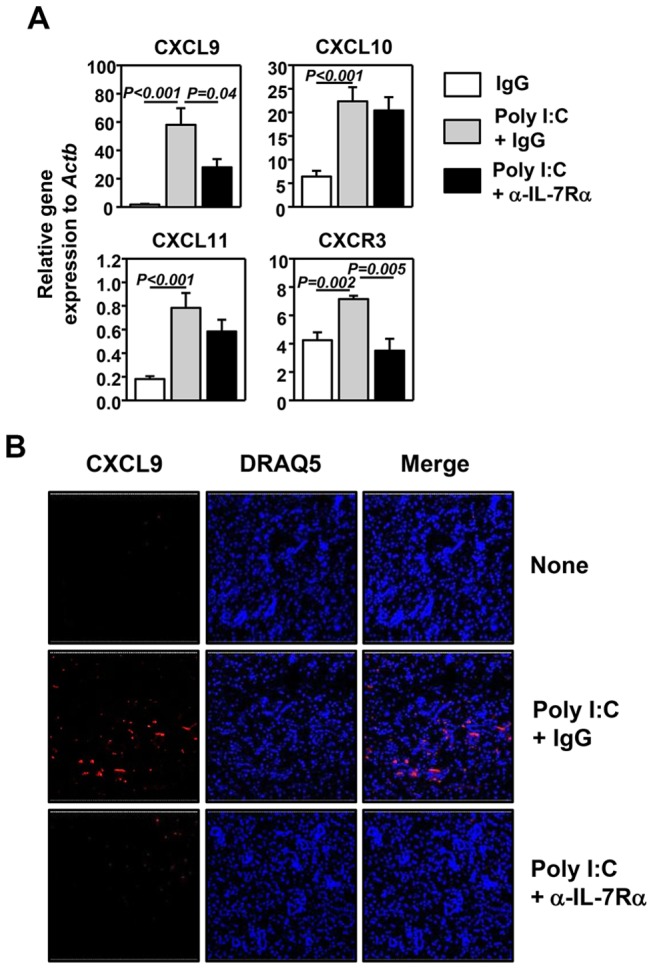
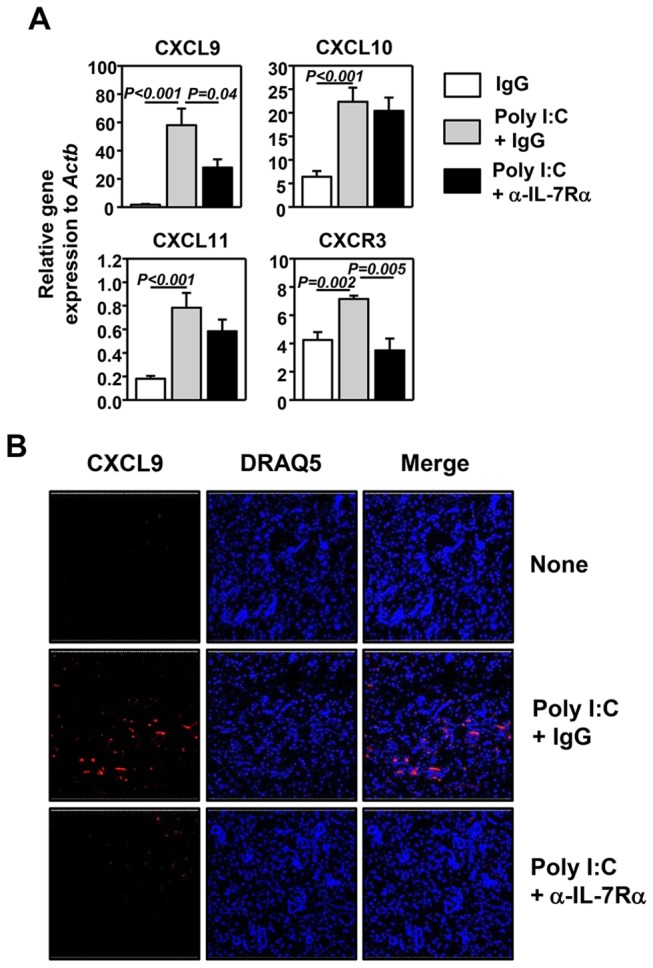
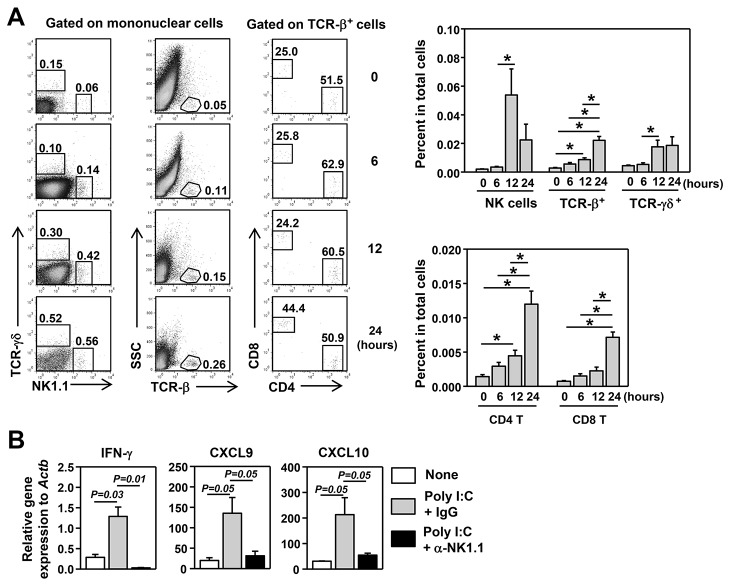
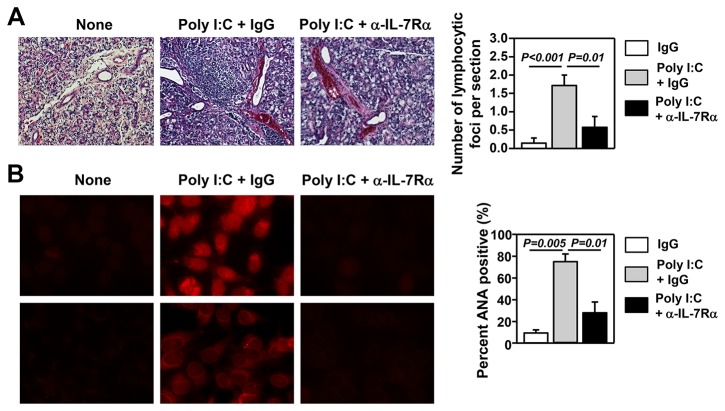
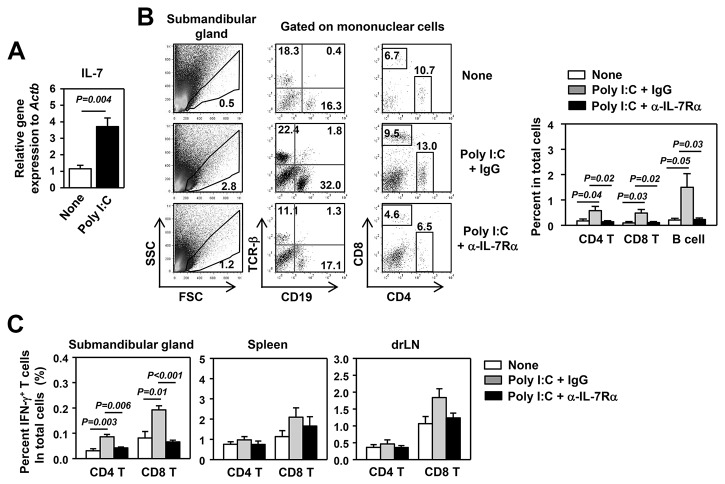
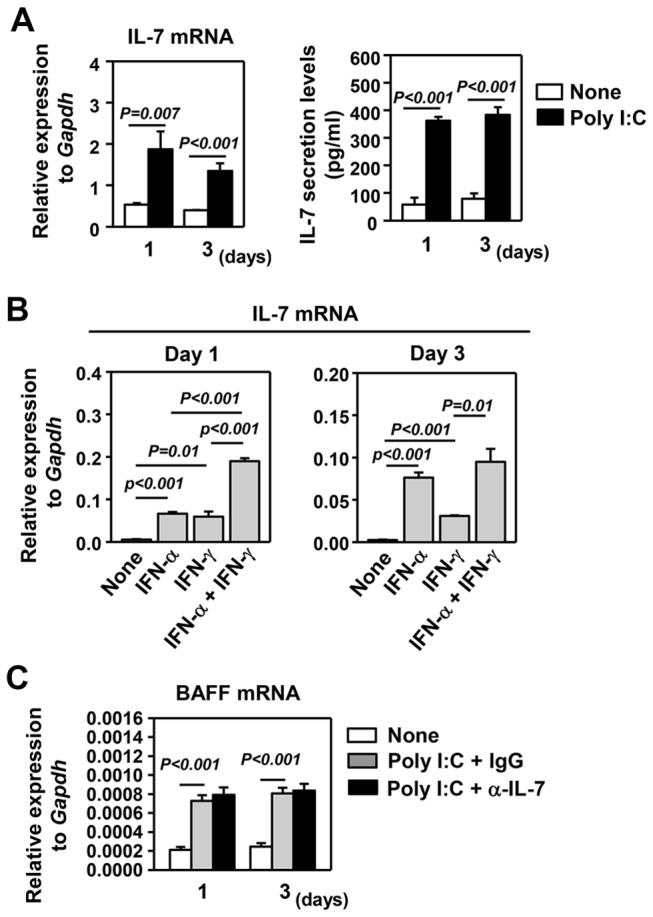
Elevated IL-7 in the target tissues is closely associated with multiple autoimmune disorders, including Sjögren's syndrome (SS). We recently found that IL-7 plays an essential role in the development and onset of primary SS (pSS) in C57BL/6.NOD-Aec1Aec2 mice, a well-defined mouse model of primary SS. However, environmental signals that cause excessive IL-7 production are not well-characterized. Innate immune signaling plays a critical role in shaping the adaptive immune responses including autoimmune responses. We and others have previously shown that innate immune signaling can induce IL-7 expression in lungs and intestines of C57BL/6 mice. In this study, we characterized the effects of poly I:C, a double-stranded RNA analog and toll-like receptor 3 agonist, on the induction of IL-7 expression in salivary glands and on pSS development. We showed that poly I:C administration to C57BL/6 mice rapidly induced IL-7 expression in the salivary glands in a type 1 IFN- and IFN-γ-dependent manner. Moreover, poly I:C-induced IL-7 contributed to the optimal up-regulation of CXCL9 in the salivary glands, which may subsequently promote recruitment of more IFN-γ-producing T cells. Repeated administration of poly I:C to C57BL/6.NOD-Aec1Aec2 mice accelerated the development of SS-like exocrinopathy, and this effect was abolished by the blockade of IL-7 receptor signaling with a neutralizing antibody. Finally, poly I:C or a combination of IFN-α and IFN-γ induced IL-7 gene expression and protein production in a human salivary gland epithelial cell line. Hence, we demonstrate that IL-7 expression in the salivary gland cells can be induced by poly I:C and delineate a crucial mechanism by which innate immune signals facilitate the development of pSS, which is through induction of IL-7 in the target tissues.
Conflict of interest statement
Figures
Similar articles
-
Innate Immune Signaling Induces IL-7 Production, Early Inflammatory Responses, and Sjögren's-Like Dacryoadenitis in C57BL/6 Mice.Invest Ophthalmol Vis Sci. 2015 Dec;56(13):7831-8. doi: 10.1167/iovs.15-17368. Invest Ophthalmol Vis Sci. 2015. PMID: 26658504 Free PMC article.
-
Interleukin-7 enhances the Th1 response to promote the development of Sjögren's syndrome-like autoimmune exocrinopathy in mice.Arthritis Rheum. 2013 Aug;65(8):2132-42. doi: 10.1002/art.38007. Arthritis Rheum. 2013. PMID: 23666710 Free PMC article.
-
Increased expression of interferon-λ in minor salivary glands of patients with primary Sjögren's syndrome and its synergic effect with interferon-α on salivary gland epithelial cells.Clin Exp Rheumatol. 2018 May-Jun;36 Suppl 112(3):31-40. Epub 2017 Apr 18. Clin Exp Rheumatol. 2018. PMID: 28421993
-
Th17 cells in primary Sjögren's syndrome: Pathogenicity and plasticity.J Autoimmun. 2018 Feb;87:16-25. doi: 10.1016/j.jaut.2017.11.003. Epub 2017 Nov 27. J Autoimmun. 2018. PMID: 29191572 Review.
-
Advances in cellular and molecular pathways of salivary gland damage in Sjögren's syndrome.Front Immunol. 2024 Jul 10;15:1405126. doi: 10.3389/fimmu.2024.1405126. eCollection 2024. Front Immunol. 2024. PMID: 39050857 Free PMC article. Review.
Cited by
-
Innate Immune Signaling Induces IL-7 Production, Early Inflammatory Responses, and Sjögren's-Like Dacryoadenitis in C57BL/6 Mice.Invest Ophthalmol Vis Sci. 2015 Dec;56(13):7831-8. doi: 10.1167/iovs.15-17368. Invest Ophthalmol Vis Sci. 2015. PMID: 26658504 Free PMC article.
-
The value of animal models to study immunopathology of primary human Sjögren's syndrome symptoms.Expert Rev Clin Immunol. 2014 Apr;10(4):469-81. doi: 10.1586/1744666X.2014.883920. Epub 2014 Feb 10. Expert Rev Clin Immunol. 2014. PMID: 24506531 Free PMC article. Review.
-
Role of JAK-STAT signaling pathway in pathogenesis and treatment of primary Sjögren's syndrome.Chin Med J (Engl). 2023 Oct 5;136(19):2297-2306. doi: 10.1097/CM9.0000000000002539. Chin Med J (Engl). 2023. PMID: 37185152 Free PMC article. Review.
-
Mitochondrial double-stranded RNAs as a pivotal mediator in the pathogenesis of Sjӧgren's syndrome.Mol Ther Nucleic Acids. 2022 Sep 27;30:257-269. doi: 10.1016/j.omtn.2022.09.020. eCollection 2022 Dec 13. Mol Ther Nucleic Acids. 2022. PMID: 36284513 Free PMC article.
-
A gene network regulated by the transcription factor VGLL3 as a promoter of sex-biased autoimmune diseases.Nat Immunol. 2017 Feb;18(2):152-160. doi: 10.1038/ni.3643. Epub 2016 Dec 19. Nat Immunol. 2017. PMID: 27992404 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
