

Engineering synthetic vaccines using cues from natural immunity
- PMID: 24150416
- PMCID: PMC3928825
- DOI: 10.1038/nmat3775
Engineering synthetic vaccines using cues from natural immunity
Abstract
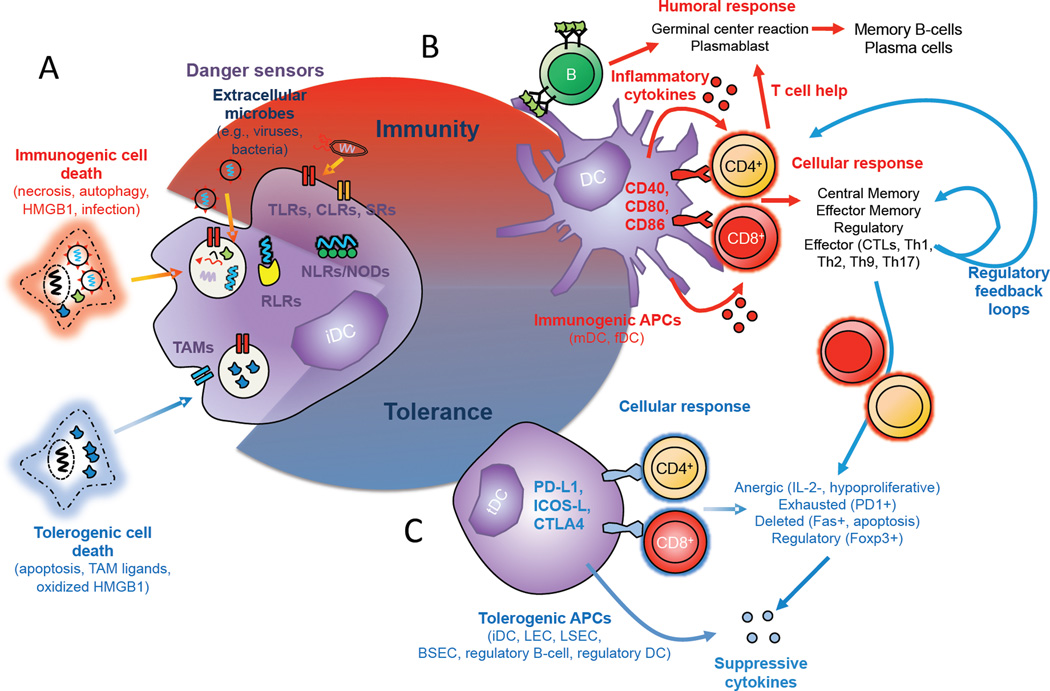
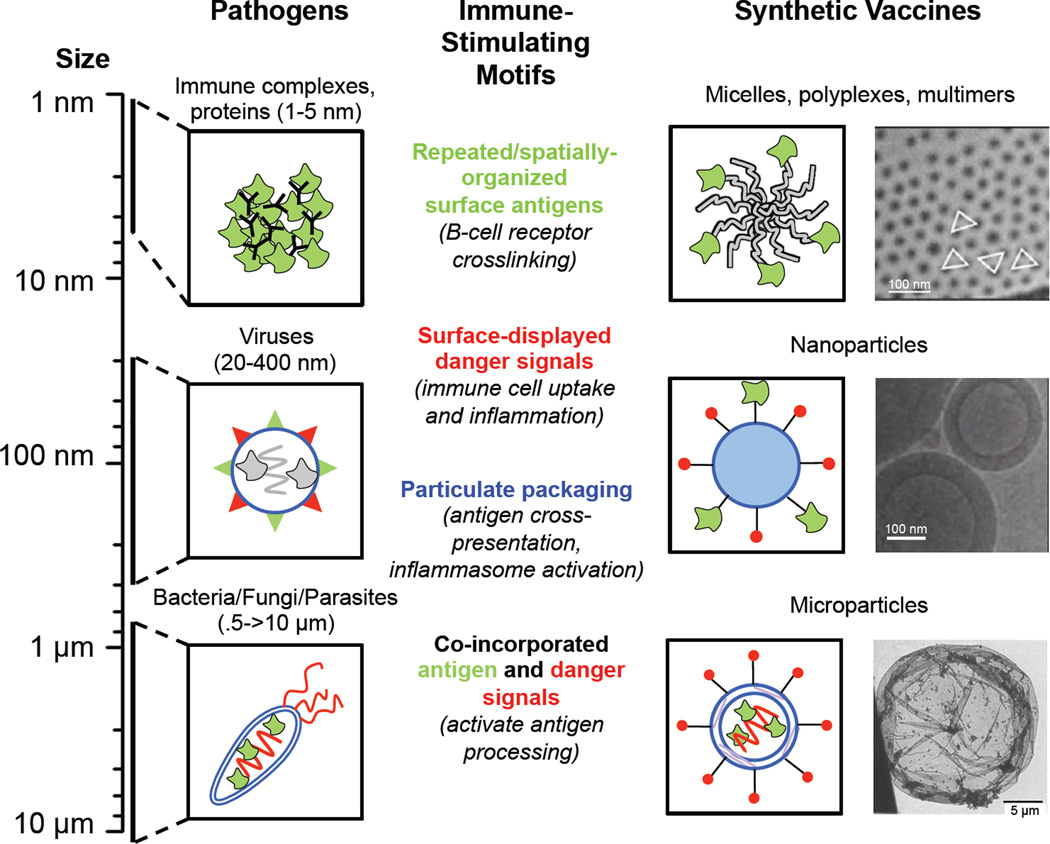
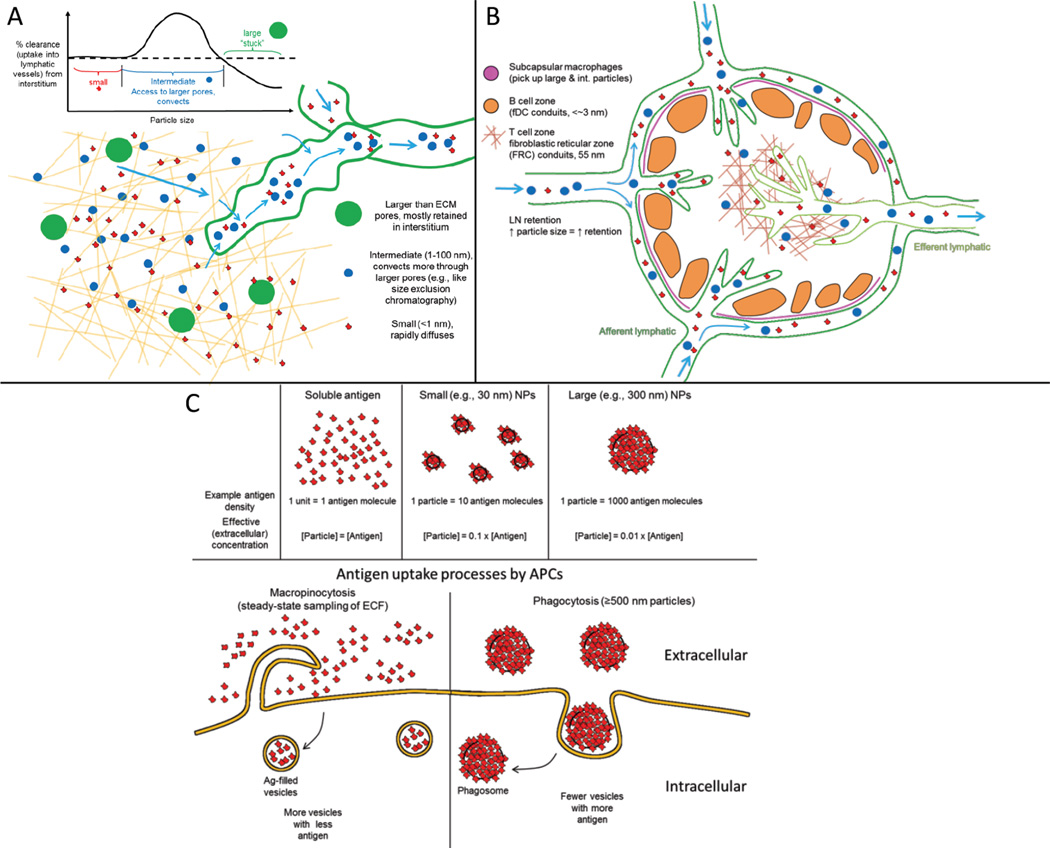
Vaccines aim to protect against or treat diseases through manipulation of the immune response, promoting either immunity or tolerance. In the former case, vaccines generate antibodies and T cells poised to protect against future pathogen encounter or attack diseased cells such as tumours; in the latter case, which is far less developed, vaccines block pathogenic autoreactive T cells and autoantibodies that target self tissue. Enormous challenges remain, however, as a consequence of our incomplete understanding of human immunity. A rapidly growing field of research is the design of vaccines based on synthetic materials to target organs, tissues, cells or intracellular compartments; to co-deliver immunomodulatory signals that control the quality of the immune response; or to act directly as immune regulators. There exists great potential for well-defined materials to further our understanding of immunity. Here we describe recent advances in the design of synthetic materials to direct immune responses, highlighting successes and challenges in prophylactic, therapeutic and tolerance-inducing vaccines.
Figures
Similar articles
-
Pathogen-like particles: biomimetic vaccine carriers engineered at the nanoscale.Curr Opin Biotechnol. 2014 Aug;28:51-8. doi: 10.1016/j.copbio.2013.11.005. Epub 2013 Dec 7. Curr Opin Biotechnol. 2014. PMID: 24832075 Review.
-
Synthetic vaccines: the role of adjuvants in immune targeting.Curr Med Chem. 2003 Aug;10(15):1423-39. doi: 10.2174/0929867033457340. Curr Med Chem. 2003. PMID: 12871139 Review.
-
The development of synthetic antitumour vaccines from mucin glycopeptide antigens.Chem Soc Rev. 2013 May 21;42(10):4421-42. doi: 10.1039/c3cs35470a. Epub 2013 Feb 25. Chem Soc Rev. 2013. PMID: 23440054 Review.
-
Synthetic peptides as vaccines.Biotechnology. 1992;20:451-65. doi: 10.1016/b978-0-7506-9265-6.50026-9. Biotechnology. 1992. PMID: 1600388 Review.
-
Recombinant Newcastle disease virus-vectored vaccines against human and animal infectious diseases.Future Microbiol. 2015;10(8):1307-23. doi: 10.2217/FMB.15.59. Epub 2015 Aug 3. Future Microbiol. 2015. PMID: 26234909 Review.
Cited by
-
Monophosphoryl lipid A-adjuvanted nucleoprotein-neuraminidase nanoparticles improve immune protection against divergent influenza viruses.Nanomedicine. 2023 Jan;47:102614. doi: 10.1016/j.nano.2022.102614. Epub 2022 Oct 17. Nanomedicine. 2023. PMID: 36265560 Free PMC article.
-
Overcoming transport barriers for interstitial-, lymphatic-, and lymph node-targeted drug delivery.Curr Opin Chem Eng. 2015 Feb 1;7:65-74. doi: 10.1016/j.coche.2014.11.003. Curr Opin Chem Eng. 2015. PMID: 25745594 Free PMC article.
-
A biomimetic yeast shell vaccine coated with layered double hydroxides induces a robust humoral and cellular immune response against tumors.Nanoscale Adv. 2020 Jun 26;2(8):3494-3506. doi: 10.1039/d0na00249f. eCollection 2020 Aug 11. Nanoscale Adv. 2020. PMID: 36134256 Free PMC article.
-
A facile approach for development of a vaccine made of bacterial double-layered membrane vesicles (DMVs).Biomaterials. 2018 Dec;187:28-38. doi: 10.1016/j.biomaterials.2018.09.042. Epub 2018 Oct 1. Biomaterials. 2018. PMID: 30292939 Free PMC article.
-
Engineering, on-demand manufacturing, and scaling-up of polymeric nanocapsules.Bioeng Transl Med. 2018 Oct 26;4(1):38-50. doi: 10.1002/btm2.10118. eCollection 2019 Jan. Bioeng Transl Med. 2018. PMID: 30680317 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
