

Colon Macrophages Polarized by Commensal Bacteria Cause Colitis and Cancer through the Bystander Effect
- PMID: 24151540
- PMCID: PMC3799201
- DOI: 10.1593/tlo.13412
Colon Macrophages Polarized by Commensal Bacteria Cause Colitis and Cancer through the Bystander Effect
Abstract
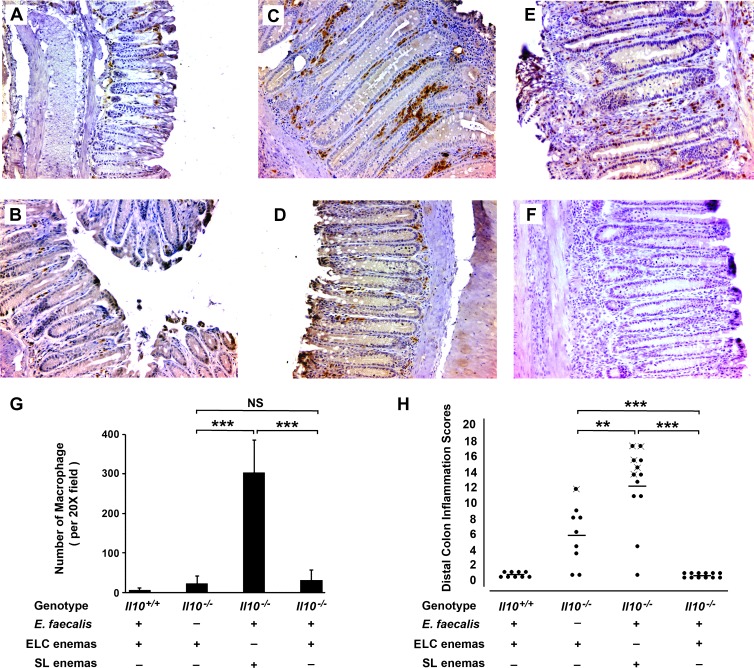
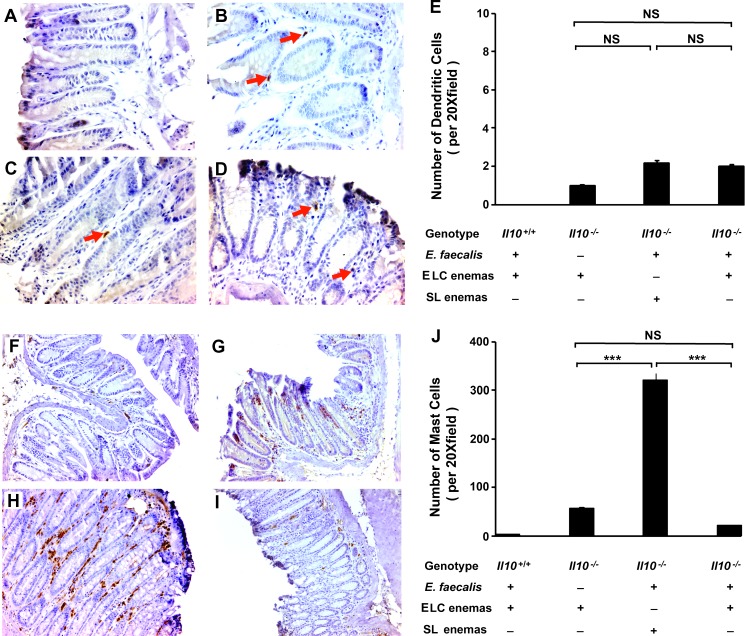
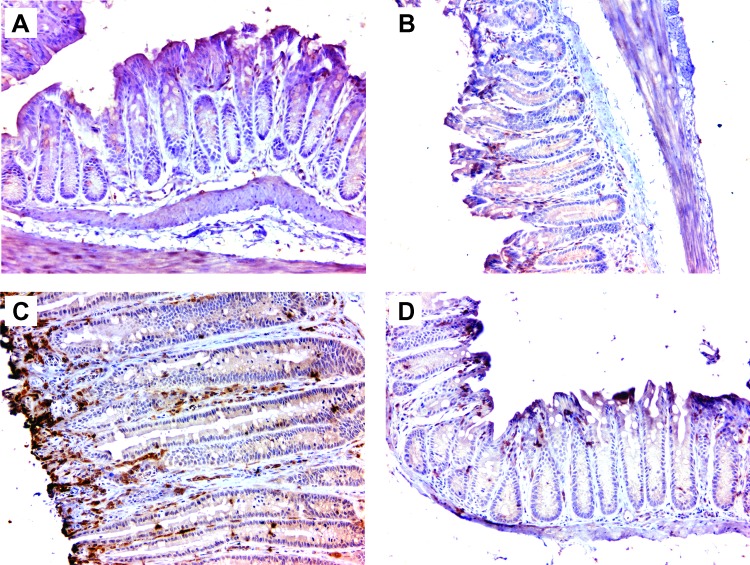
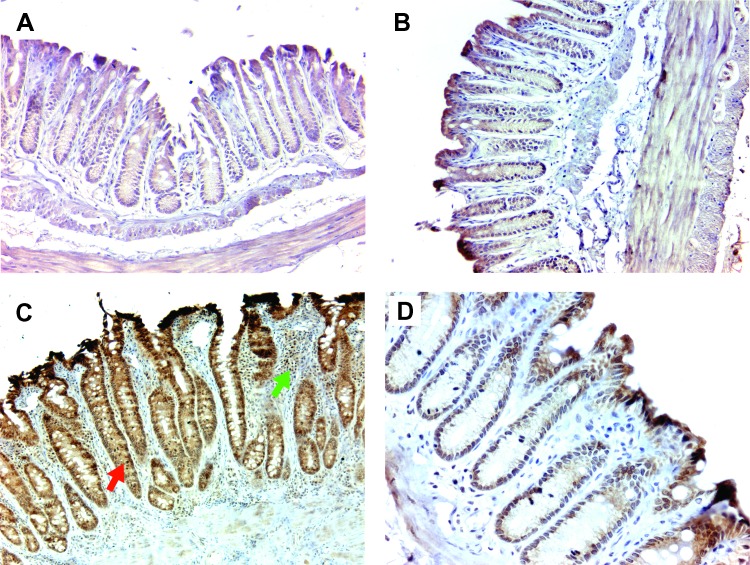
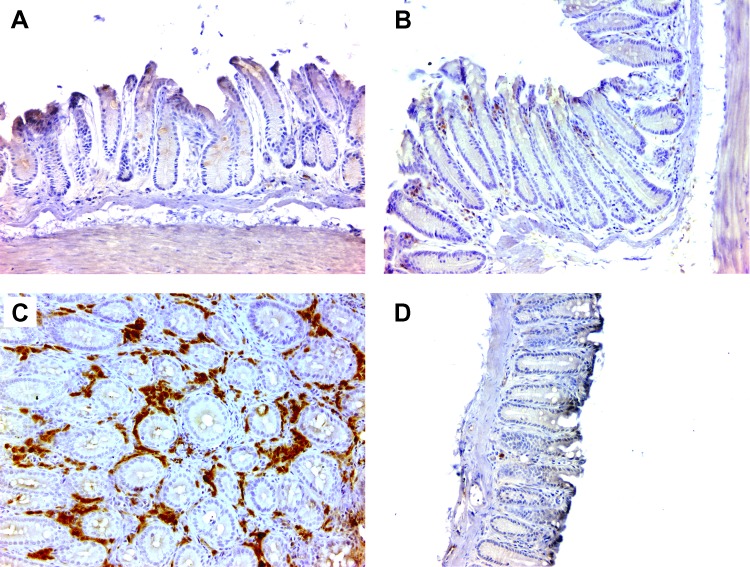
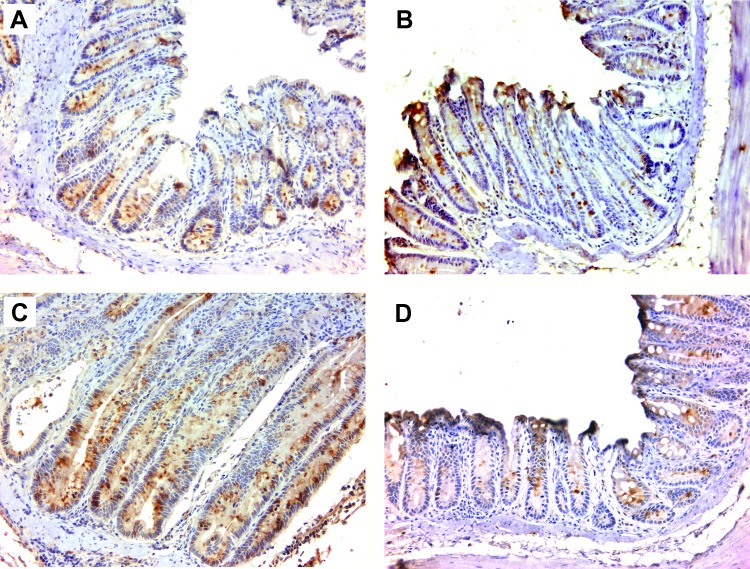
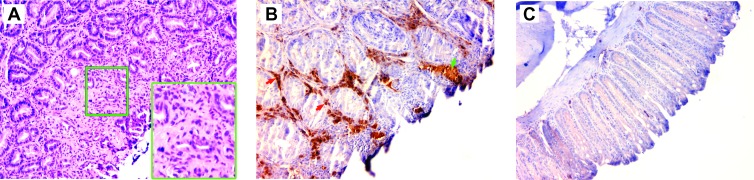
Intestinal commensal bacteria have recently been shown to trigger macrophages to produce diffusible clastogens (or chromosome-breaking factors) through a bystander effect (BSE) that mediates DNA damage and induces chromosomal instability in neighboring cells. Colon macrophages appear central to colon carcinogenesis and BSE through the expression of tumor necrosis factor-α (TNF-α) and cyclooxygenase-2 (COX-2). The former induces netrin-1, a regulator of intestinal epithelial cell apoptosis, and the latter generates trans-4-hydroxy-2-nonenal (4-HNE), an endogenous mutagen. To test whether colon macrophages are key effectors for BSE, we depleted these cells in interleukin-10 knockout mice colonized with Enterococcus faecalis using encapsulated liposomal clodronate (ELC), a bisphosphonate that causes macrophage apoptosis. We observed that E. faecalis polarizes colon macrophages to an M1 phenotype. In addition, depleting these cells suppressed COX-2 and TNF-α, blocked the formation of 4-HNE protein adducts, and inhibited up-regulation of netrin-1-all markers for BSE. Finally, treatment with ELC prevented colitis, β-catenin activation, and cancer formation. These results show that selected human commensals can polarize colon macrophages to the M1 phenotype and, when activated, serve as the key effector for bacterial-induced BSE. Our findings suggest that depleting M1-polarized macro-phages is a mechanism for the chemopreventive activity of bisphosphonates and that it represents a new strategy for preventing colon cancer induced by intestinal commensals.
Figures
References
-
- Colotta F, Allavena P, Sica A, Garlanda C, Mantovani A. Cancer-related inflammation, the seventh hallmark of cancer: links to genetic instability. Carcinogenesis. 2009;30:1073–1081. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials