

The Rac-GAP Bcr is a novel regulator of the Par complex that controls cell polarity
- PMID: 24152735
- PMCID: PMC3861082
- DOI: 10.1091/mbc.E13-06-0333
The Rac-GAP Bcr is a novel regulator of the Par complex that controls cell polarity
Abstract
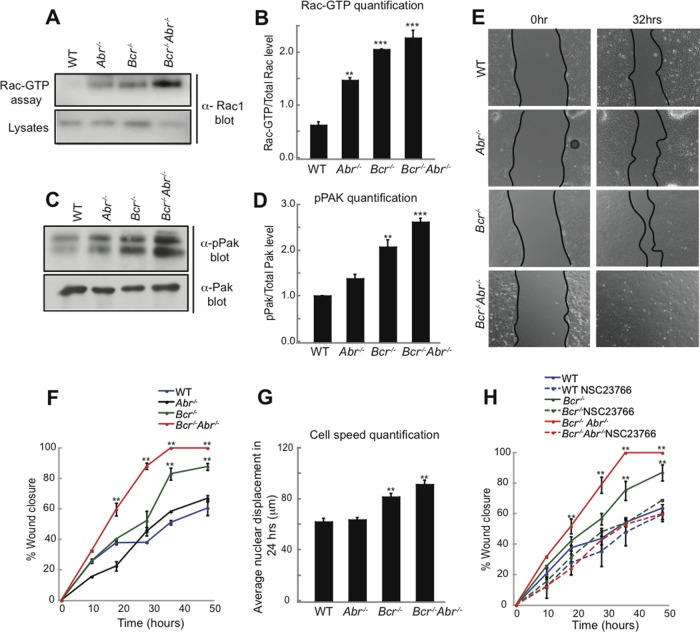
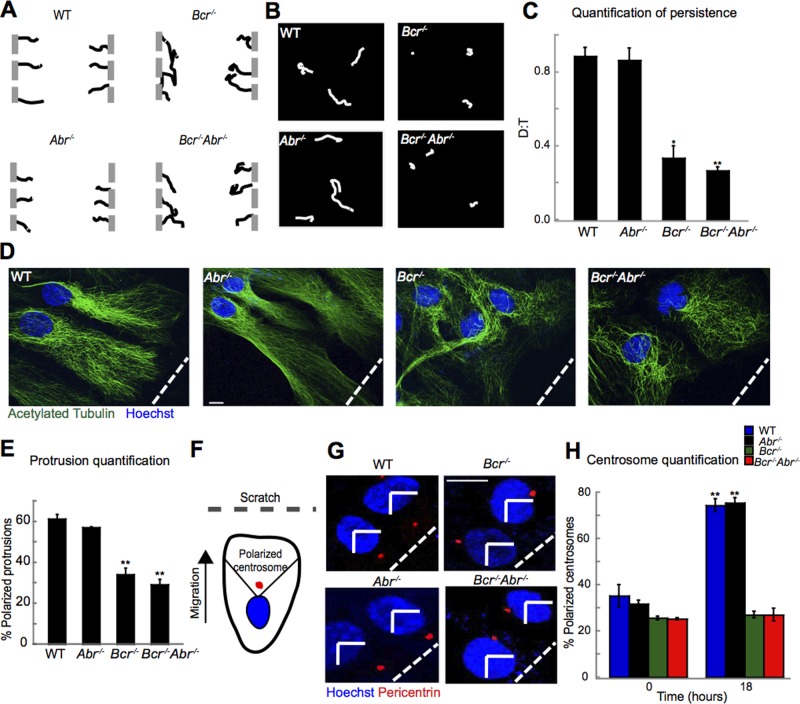
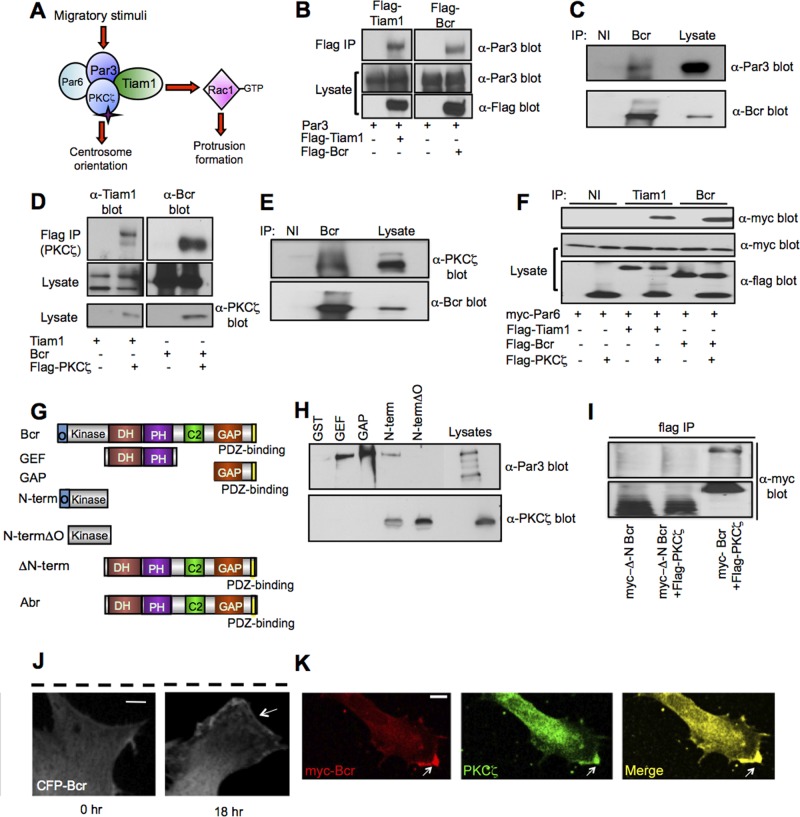
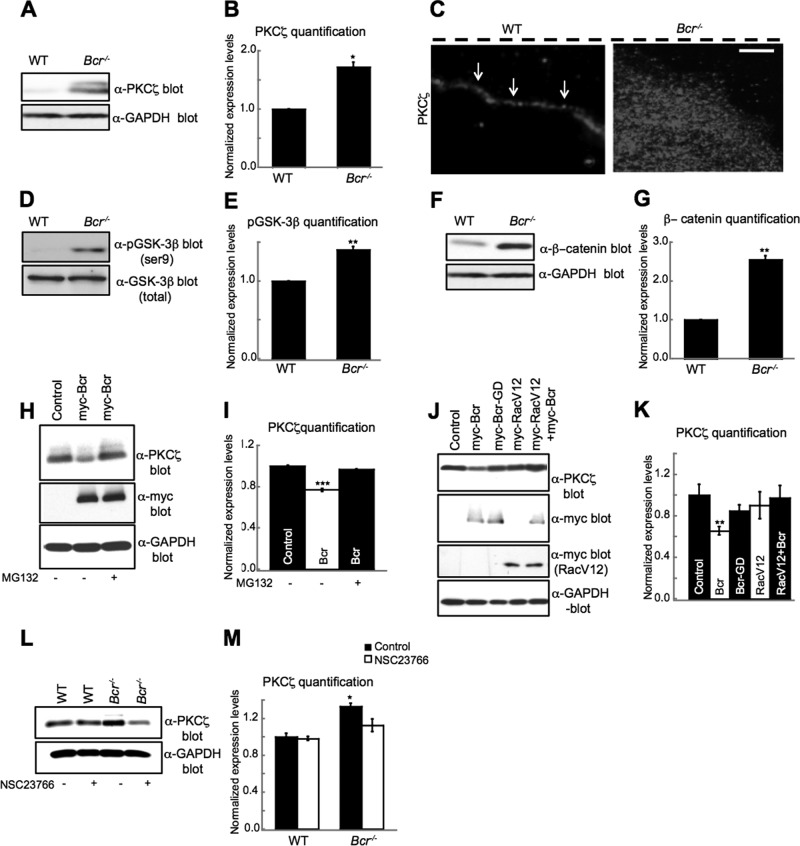
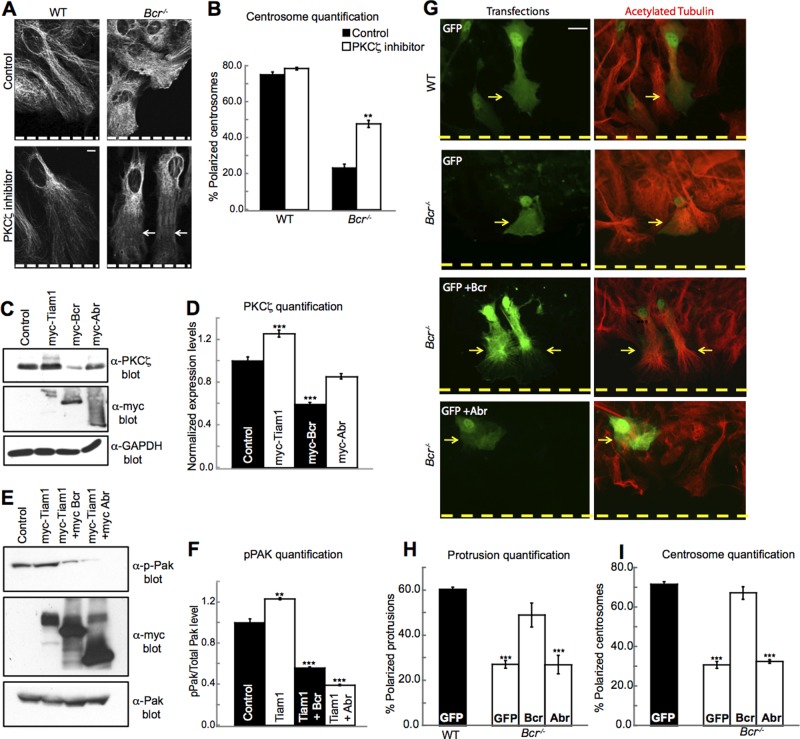
Cell polarization is essential for many biological processes, including directed cell migration, and loss of polarity contributes to pathological conditions such as cancer. The Par complex (Par3, Par6, and PKCζ) controls cell polarity in part by recruiting the Rac-specific guanine nucleotide exchange factor T-lymphoma invasion and metastasis 1 (Tiam1) to specialized cellular sites, where Tiam1 promotes local Rac1 activation and cytoskeletal remodeling. However, the mechanisms that restrict Par-Tiam1 complex activity to the leading edge to maintain cell polarity during migration remain unclear. We identify the Rac-specific GTPase-activating protein (GAP) breakpoint cluster region protein (Bcr) as a novel regulator of the Par-Tiam1 complex. We show that Bcr interacts with members of the Par complex and inhibits both Rac1 and PKCζ signaling. Loss of Bcr results in faster, more random migration and striking polarity defects in astrocytes. These polarity defects are rescued by reducing PKCζ activity or by expressing full-length Bcr, but not an N-terminal deletion mutant or the homologous Rac-GAP, Abr, both of which fail to associate with the Par complex. These results demonstrate that Bcr is an integral member of the Par-Tiam1 complex that controls polarized cell migration by locally restricting both Rac1 and PKCζ function.
Figures
References
-
- Cau J, Hall A. Cdc42 controls the polarity of the actin and microtubule cytoskeletons through two distinct signal transduction pathways. J Cell Sci. 2005;118:2579–2587. - PubMed
-
- Chan JR, Jolicoeur C, Yamauchi J, Elliott J, Fawcett JP, Ng BK, Cayouette M. The polarity protein Par-3 directly interacts with p75NTR to regulate myelination. Science. 2006;314:832–836. - PubMed
-
- Chen X, Macara IG. Par-3 controls tight junction assembly through the Rac exchange factor Tiam1. Nat Cell Biol. 2005;7:262–269. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
