

A diurnal serum lipid integrates hepatic lipogenesis and peripheral fatty acid use
- PMID: 24153306
- PMCID: PMC4141623
- DOI: 10.1038/nature12710
A diurnal serum lipid integrates hepatic lipogenesis and peripheral fatty acid use
Abstract
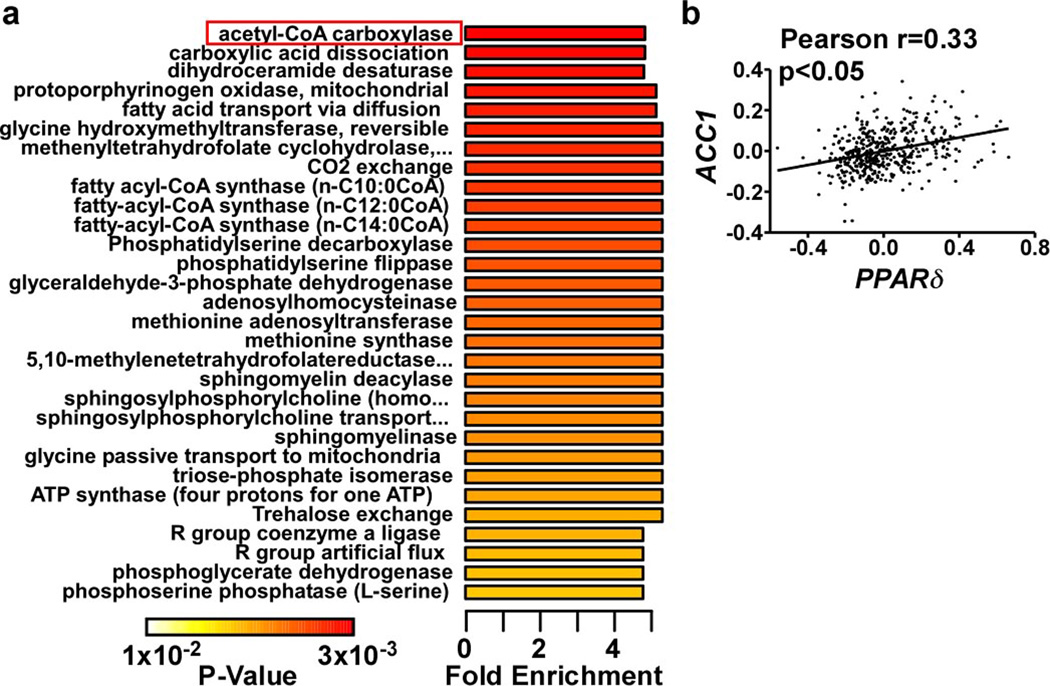
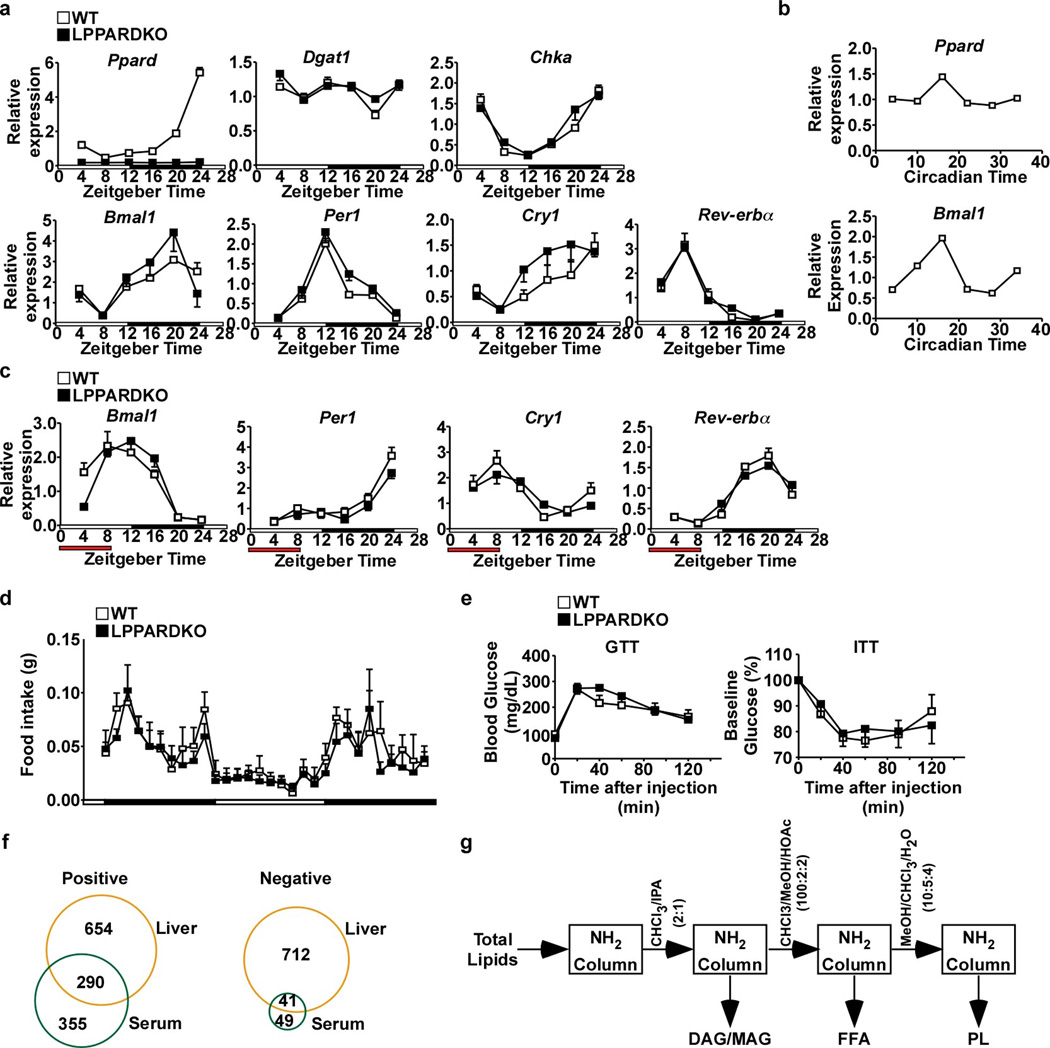
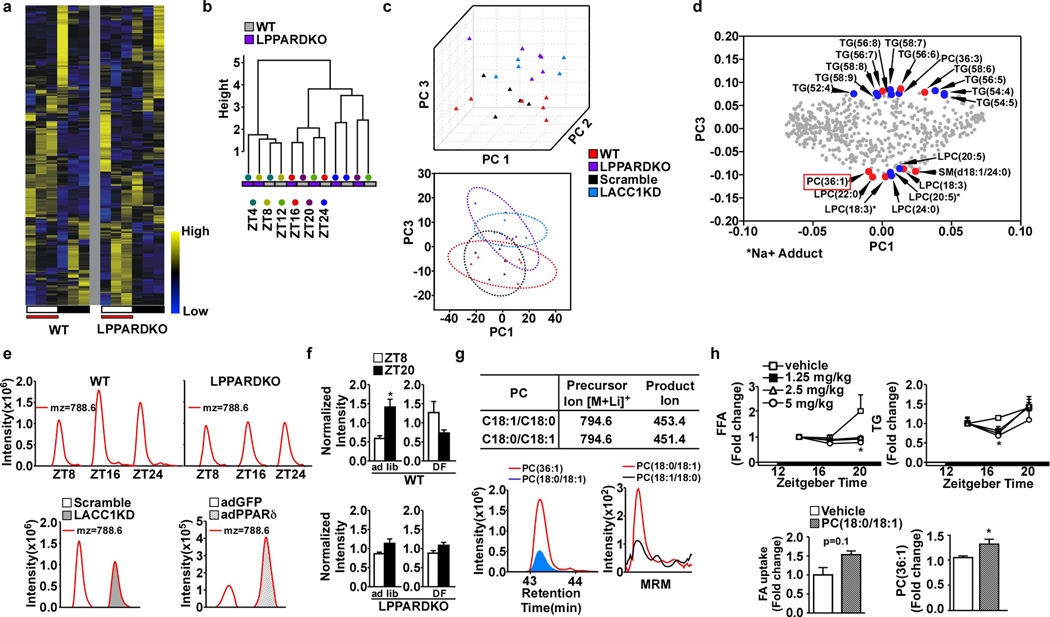
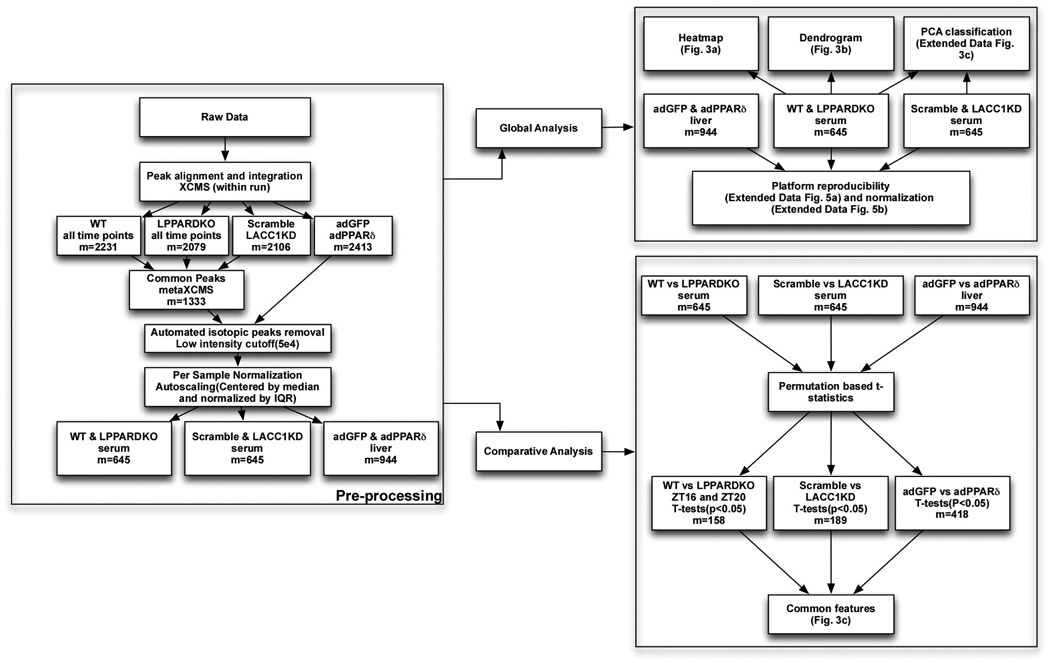
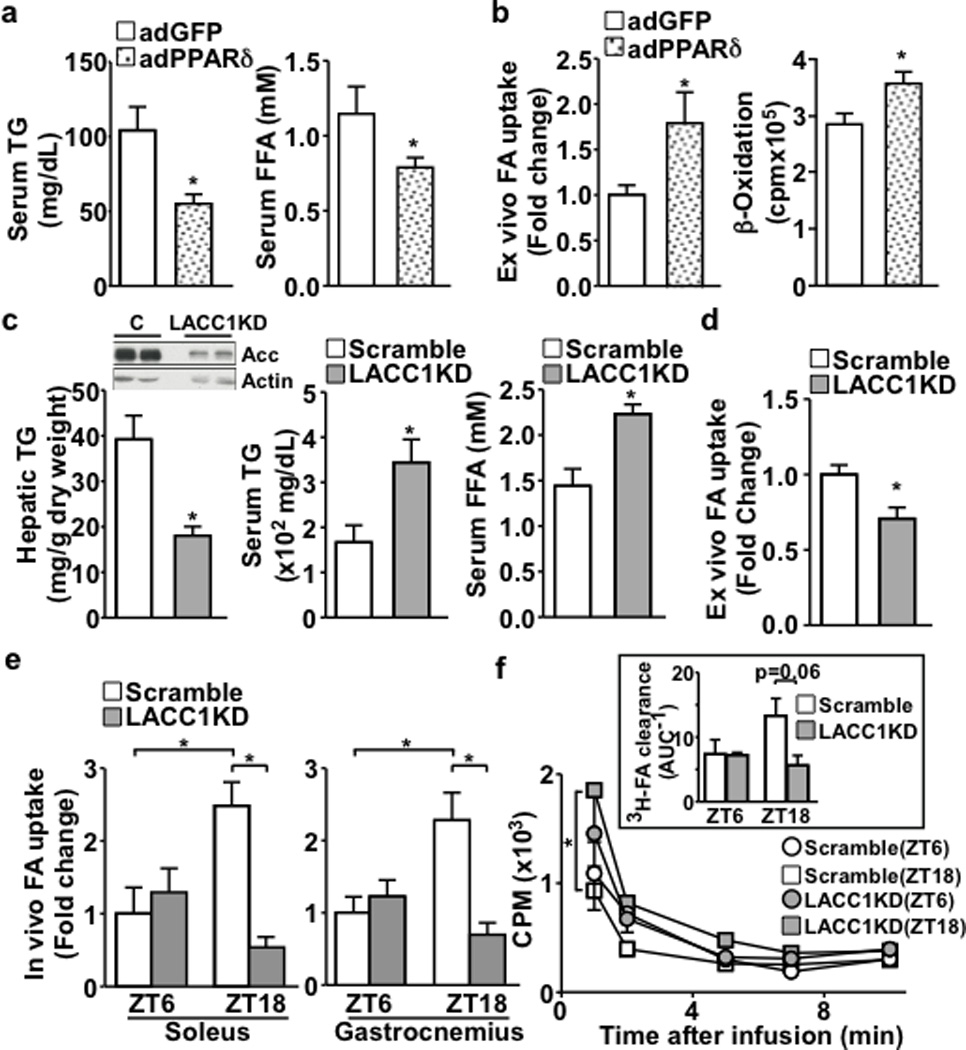
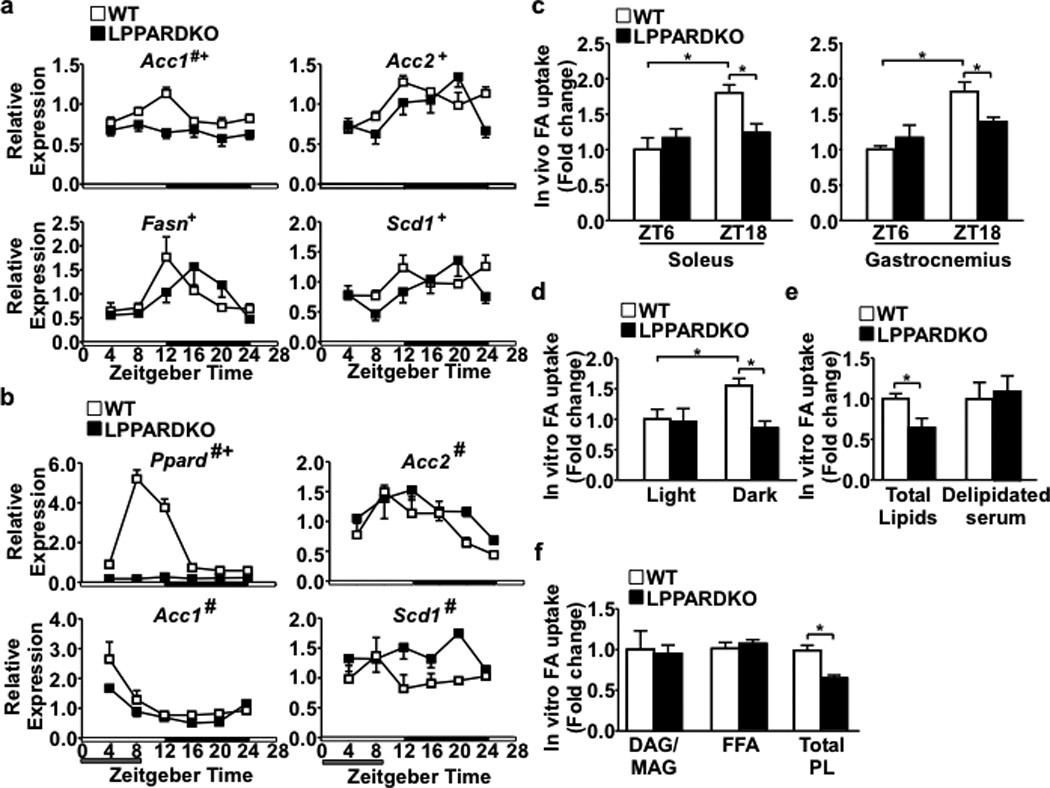
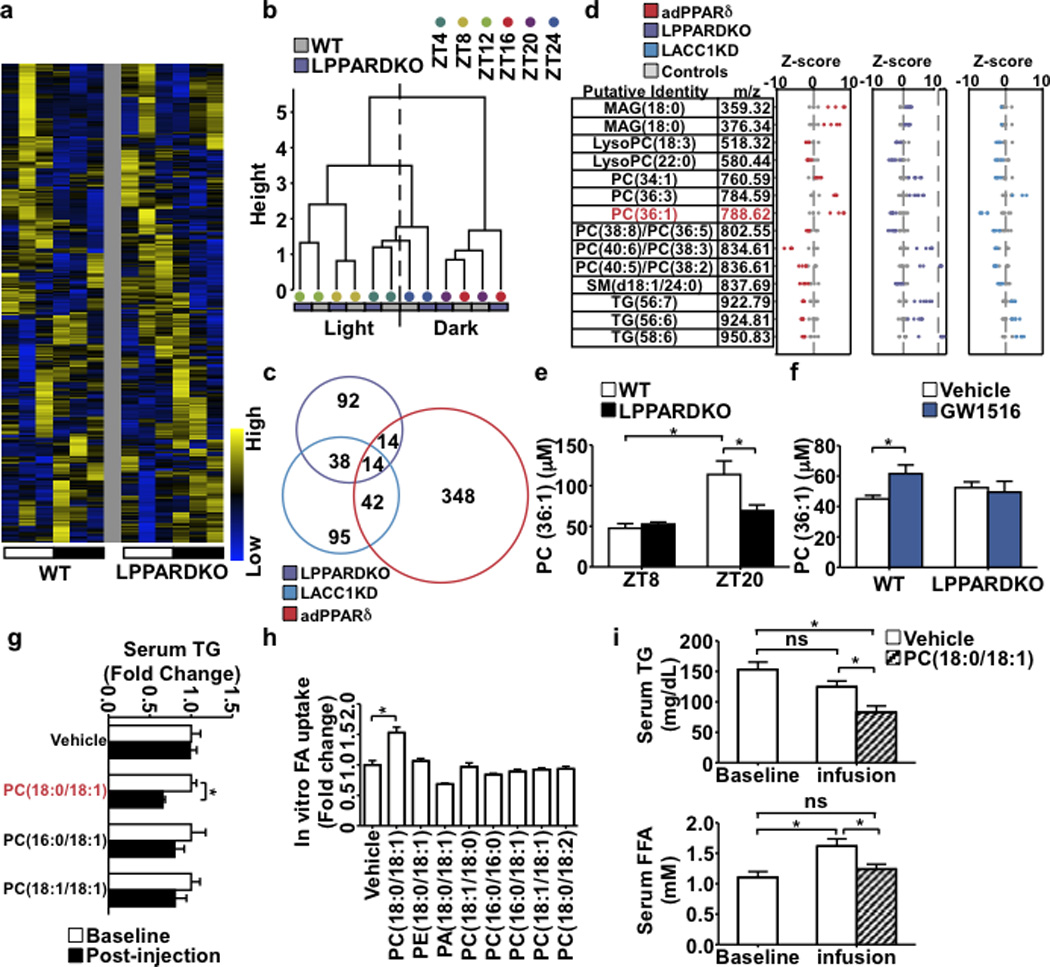
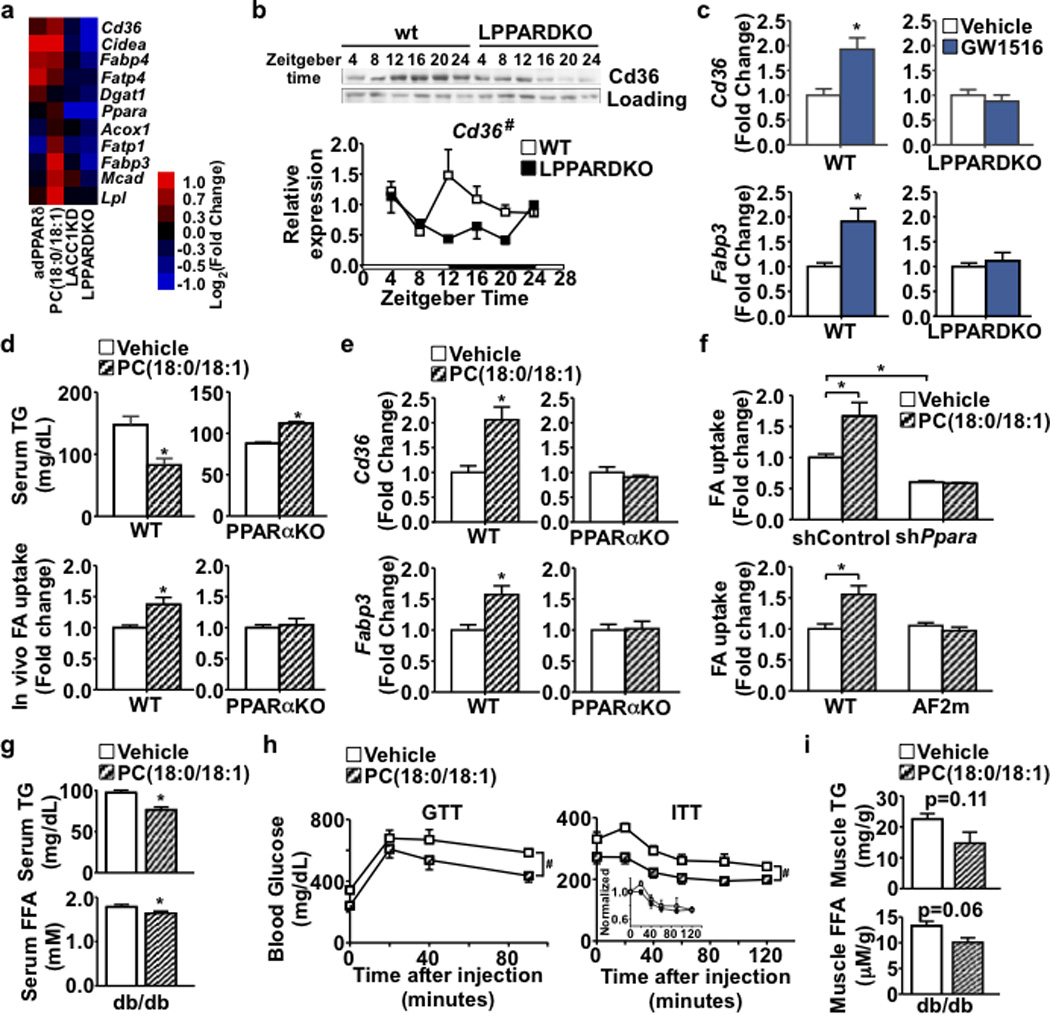
Food intake increases the activity of hepatic de novo lipogenesis, which mediates the conversion of glucose to fats for storage or use. In mice, this program follows a circadian rhythm that peaks with nocturnal feeding and is repressed by Rev-erbα/β and an HDAC3-containing complex during the day. The transcriptional activators controlling rhythmic lipid synthesis in the dark cycle remain poorly defined. Disturbances in hepatic lipogenesis are also associated with systemic metabolic phenotypes, suggesting that lipogenesis in the liver communicates with peripheral tissues to control energy substrate homeostasis. Here we identify a PPARδ-dependent de novo lipogenic pathway in the liver that modulates fat use by muscle via a circulating lipid. The nuclear receptor PPARδ controls diurnal expression of lipogenic genes in the dark/feeding cycle. Liver-specific PPARδ activation increases, whereas hepatocyte-Ppard deletion reduces, muscle fatty acid uptake. Unbiased metabolite profiling identifies phosphatidylcholine 18:0/18:1 (PC(18:0/18:1) as a serum lipid regulated by diurnal hepatic PPARδ activity. PC(18:0/18:1) reduces postprandial lipid levels and increases fatty acid use through muscle PPARα. High-fat feeding diminishes rhythmic production of PC(18:0/18:1), whereas PC(18:0/18:1) administration in db/db mice (also known as Lepr(-/-)) improves metabolic homeostasis. These findings reveal an integrated regulatory circuit coupling lipid synthesis in the liver to energy use in muscle by coordinating the activity of two closely related nuclear receptors. These data implicate alterations in diurnal hepatic PPARδ-PC(18:0/18:1) signalling in metabolic disorders, including obesity.
Figures

Comment in
-
Physiology: A metabolic minuet.Nature. 2013 Oct 24;502(7472):454-5. doi: 10.1038/502454a. Nature. 2013. PMID: 24153294 No abstract available.
Similar articles
-
Peroxisome proliferator-activated receptor-delta induces insulin-induced gene-1 and suppresses hepatic lipogenesis in obese diabetic mice.Hepatology. 2008 Aug;48(2):432-41. doi: 10.1002/hep.22334. Hepatology. 2008. PMID: 18627005
-
Acetyl-CoA carboxylase 2-/- mutant mice are protected against fatty liver under high-fat, high-carbohydrate dietary and de novo lipogenic conditions.J Biol Chem. 2012 Apr 6;287(15):12578-88. doi: 10.1074/jbc.M111.309559. Epub 2012 Feb 23. J Biol Chem. 2012. PMID: 22362781 Free PMC article.
-
Role of peroxisome proliferator-activated receptor {delta}/{beta} in hepatic metabolic regulation.J Biol Chem. 2011 Jan 14;286(2):1237-47. doi: 10.1074/jbc.M110.138115. Epub 2010 Nov 8. J Biol Chem. 2011. PMID: 21059653 Free PMC article.
-
Circadian epigenomic remodeling and hepatic lipogenesis: lessons from HDAC3.Cold Spring Harb Symp Quant Biol. 2011;76:49-55. doi: 10.1101/sqb.2011.76.011494. Epub 2011 Sep 6. Cold Spring Harb Symp Quant Biol. 2011. PMID: 21900149 Free PMC article. Review.
-
Hepatic lipid accumulation: cause and consequence of dysregulated glucoregulatory hormones.J Endocrinol. 2017 Jul;234(1):R1-R21. doi: 10.1530/JOE-16-0513. Epub 2017 Apr 20. J Endocrinol. 2017. PMID: 28428362 Review.
Cited by
-
The lipogenic regulator Sterol Regulatory Element Binding Factor-1c is required to maintain peripheral nerve structure and function.Springerplus. 2015 Jun 12;4(Suppl 1):L45. doi: 10.1186/2193-1801-4-S1-L45. eCollection 2015. Springerplus. 2015. PMID: 27386207 Free PMC article. No abstract available.
-
Hepatic PTEN Signaling Regulates Systemic Metabolic Homeostasis through Hepatokines-Mediated Liver-to-Peripheral Organs Crosstalk.Int J Mol Sci. 2022 Apr 2;23(7):3959. doi: 10.3390/ijms23073959. Int J Mol Sci. 2022. PMID: 35409319 Free PMC article.
-
Insights into the Role of PPARβ/δ in NAFLD.Int J Mol Sci. 2018 Jun 27;19(7):1893. doi: 10.3390/ijms19071893. Int J Mol Sci. 2018. PMID: 29954129 Free PMC article. Review.
-
The heart-liver metabolic axis: defective communication exacerbates disease.EMBO Mol Med. 2014 Apr;6(4):436-8. doi: 10.1002/emmm.201303800. Epub 2014 Mar 12. EMBO Mol Med. 2014. PMID: 24623378 Free PMC article.
-
Lipid metabolites as metabolic messengers in inter-organ communication.Trends Endocrinol Metab. 2014 Jul;25(7):356-63. doi: 10.1016/j.tem.2014.05.002. Epub 2014 Jun 2. Trends Endocrinol Metab. 2014. PMID: 24895003 Free PMC article. Review.
References
-
- Asher G, Schibler U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell metabolism. 2011;13:125–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
