

Targeting RNA foci in iPSC-derived motor neurons from ALS patients with a C9ORF72 repeat expansion
- PMID: 24154603
- PMCID: PMC4090945
- DOI: 10.1126/scitranslmed.3007529
Targeting RNA foci in iPSC-derived motor neurons from ALS patients with a C9ORF72 repeat expansion
Abstract
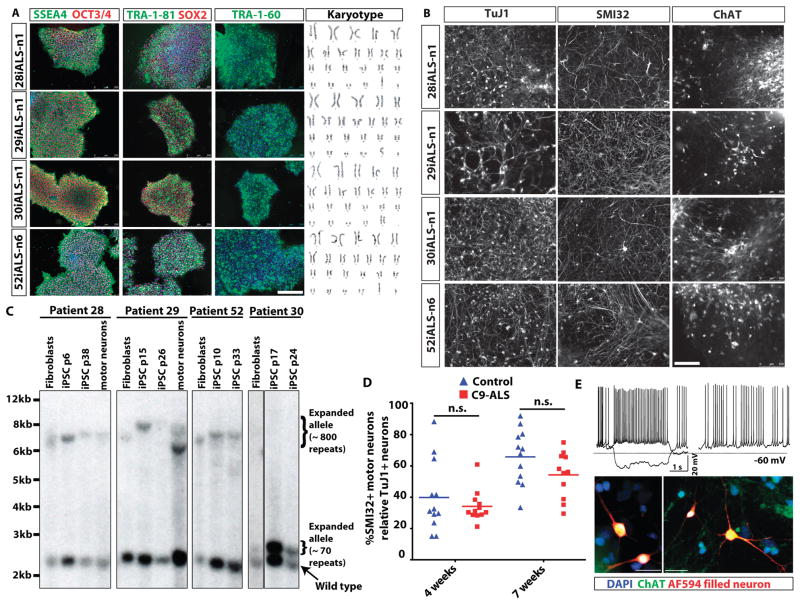
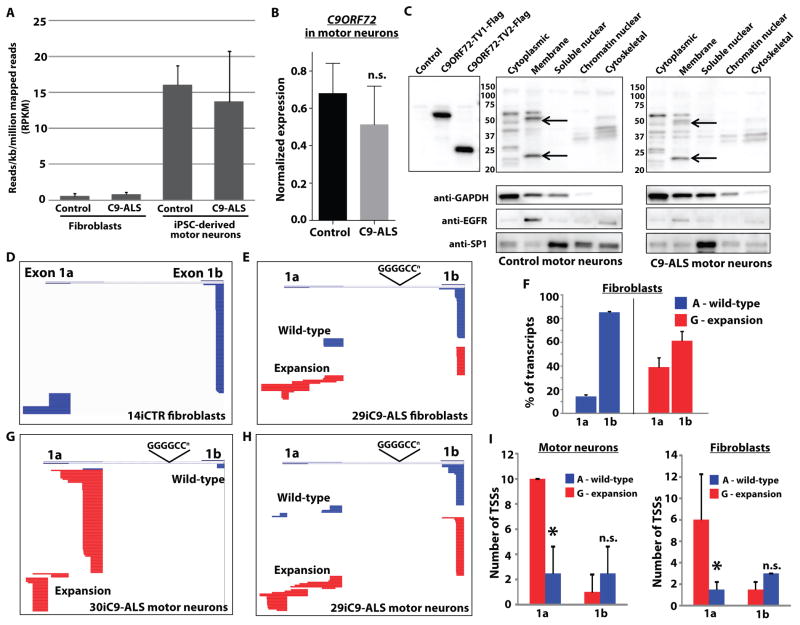
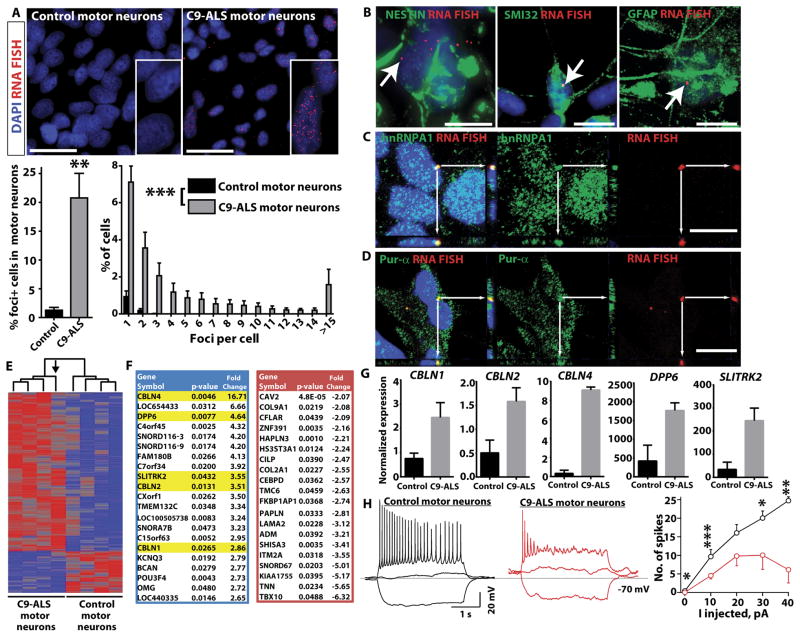
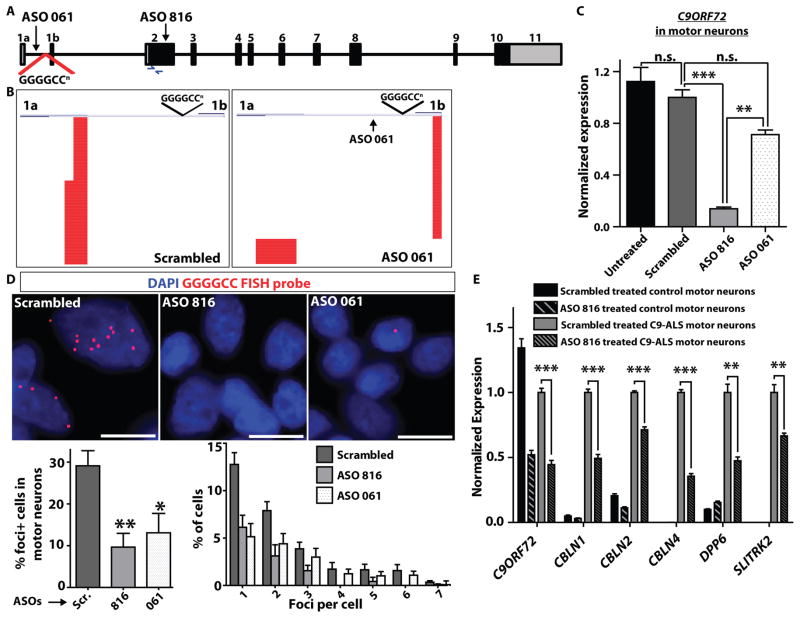
Amyotrophic lateral sclerosis (ALS) is a severe neurodegenerative condition characterized by loss of motor neurons in the brain and spinal cord. Expansions of a hexanucleotide repeat (GGGGCC) in the noncoding region of the C9ORF72 gene are the most common cause of the familial form of ALS (C9-ALS), as well as frontotemporal lobar degeneration and other neurological diseases. How the repeat expansion causes disease remains unclear, with both loss of function (haploinsufficiency) and gain of function (either toxic RNA or protein products) proposed. We report a cellular model of C9-ALS with motor neurons differentiated from induced pluripotent stem cells (iPSCs) derived from ALS patients carrying the C9ORF72 repeat expansion. No significant loss of C9ORF72 expression was observed, and knockdown of the transcript was not toxic to cultured human motor neurons. Transcription of the repeat was increased, leading to accumulation of GGGGCC repeat-containing RNA foci selectively in C9-ALS iPSC-derived motor neurons. Repeat-containing RNA foci colocalized with hnRNPA1 and Pur-α, suggesting that they may be able to alter RNA metabolism. C9-ALS motor neurons showed altered expression of genes involved in membrane excitability including DPP6, and demonstrated a diminished capacity to fire continuous spikes upon depolarization compared to control motor neurons. Antisense oligonucleotides targeting the C9ORF72 transcript suppressed RNA foci formation and reversed gene expression alterations in C9-ALS motor neurons. These data show that patient-derived motor neurons can be used to delineate pathogenic events in ALS.
Conflict of interest statement
Figures
References
-
- Lomen-Hoerth C, Anderson T, Miller B. The overlap of amyotrophic lateral sclerosis and frontotemporal dementia. Neurology. 2002 Oct 8;59:1077. - PubMed
-
- Neumann M, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006 Oct 6;314:130. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K08 NS055980/NS/NINDS NIH HHS/United States
- K08 NS075094/NS/NINDS NIH HHS/United States
- R21 NS074121/NS/NINDS NIH HHS/United States
- R01 NS069669/NS/NINDS NIH HHS/United States
- NS069669/NS/NINDS NIH HHS/United States
- NS055980/NS/NINDS NIH HHS/United States
- R01 ES20395/ES/NIEHS NIH HHS/United States
- R01 ES020395/ES/NIEHS NIH HHS/United States
- U24NS07837/NS/NINDS NIH HHS/United States
- R01 NS063964/NS/NINDS NIH HHS/United States
- RC2 NS069422/NS/NINDS NIH HHS/United States
- R21 NS084528/NS/NINDS NIH HHS/United States
- P30 AR057230/AR/NIAMS NIH HHS/United States
- R01 AG026251/AG/NIA NIH HHS/United States
- R01 NS077402/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
