

JAK1 truncating mutations in gynecologic cancer define new role of cancer-associated protein tyrosine kinase aberrations
- PMID: 24154688
- PMCID: PMC3807107
- DOI: 10.1038/srep03042
JAK1 truncating mutations in gynecologic cancer define new role of cancer-associated protein tyrosine kinase aberrations
Abstract
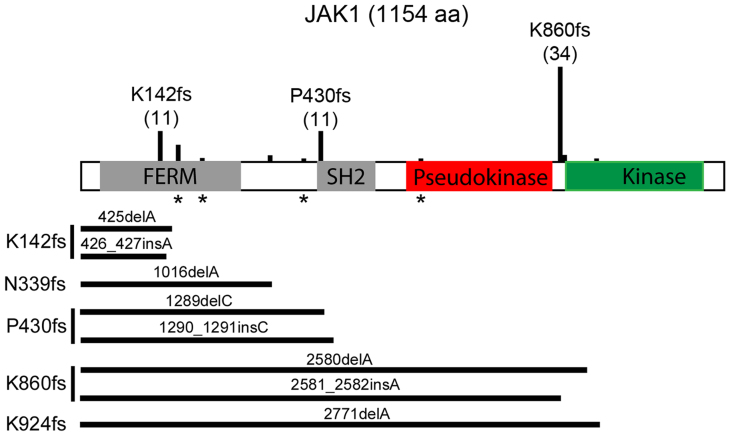
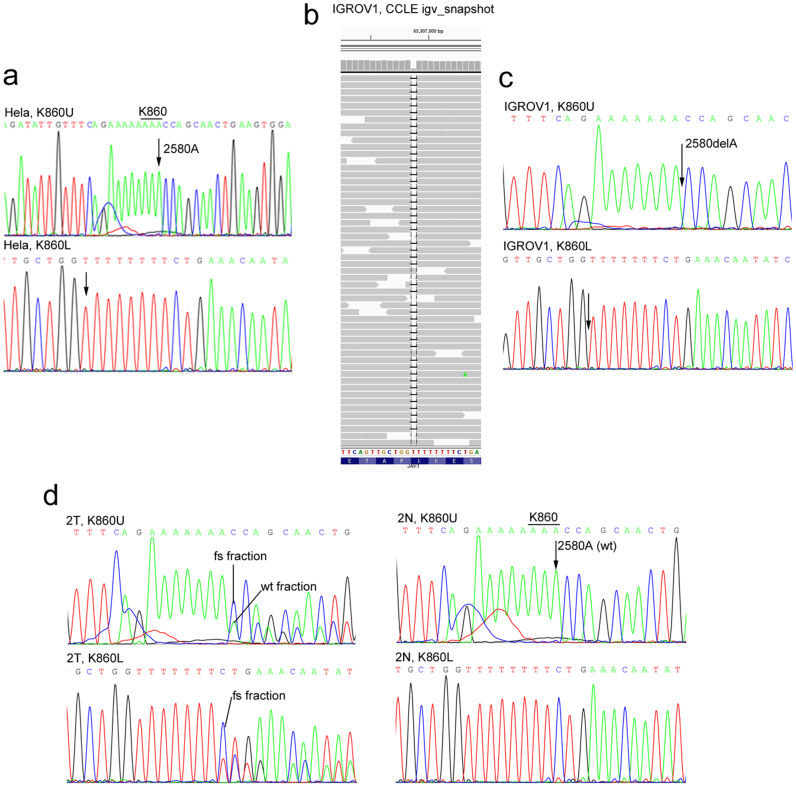
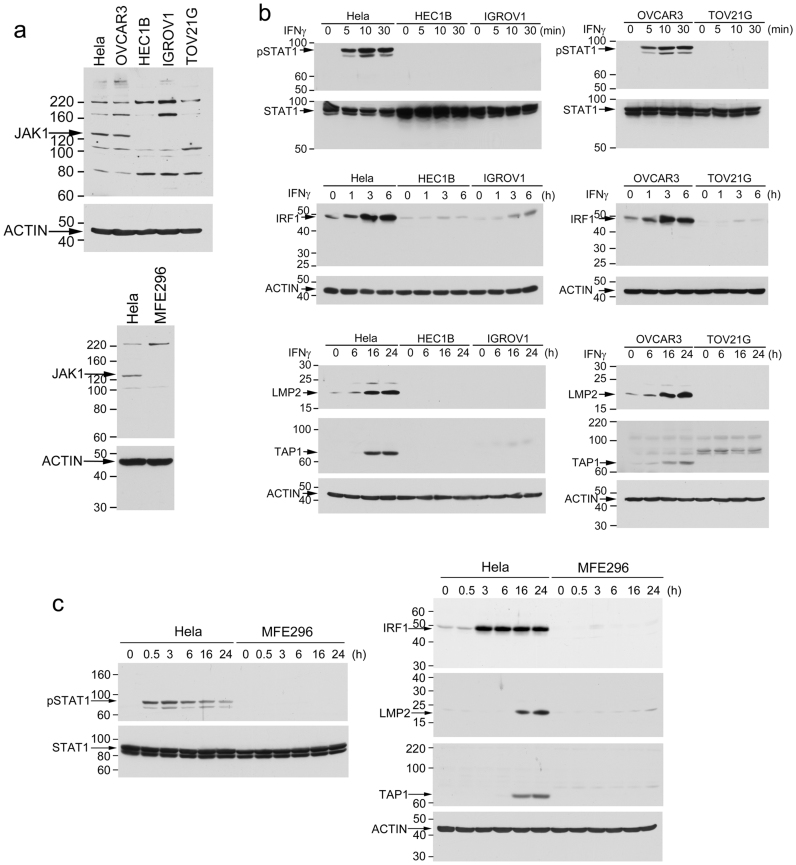
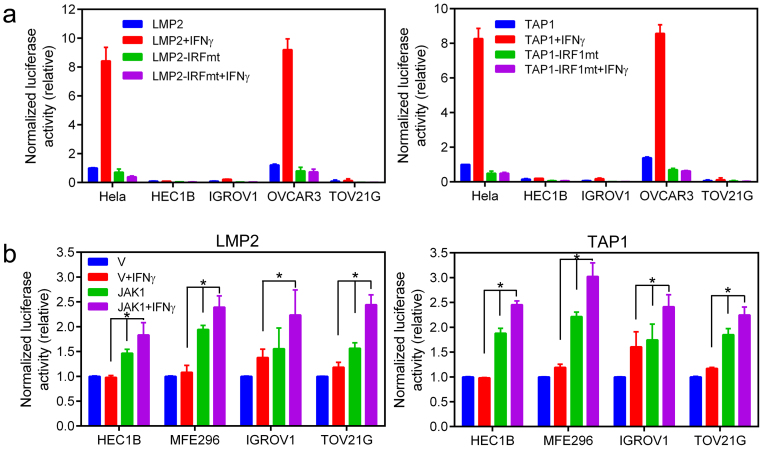
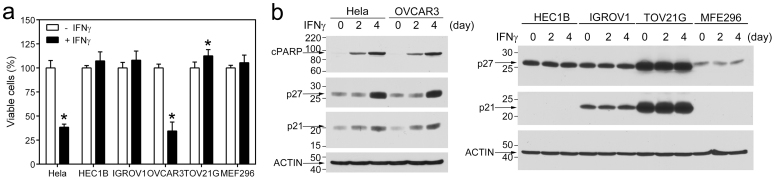
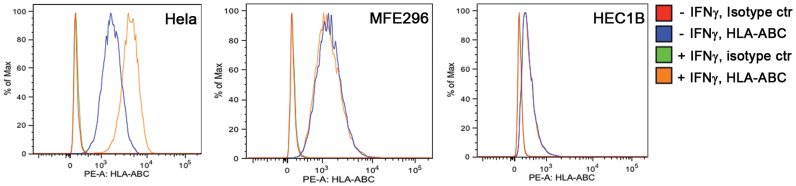
Cancer-associated protein tyrosine kinase (PTK) mutations usually are gain-of-function (GOF) mutations that drive tumor growth and metastasis. We have found 50 JAK1 truncating mutations in 36 of 635 gynecologic tumors in the Total Cancer Care® (TCC®) tumor bank. Among cancer cell lines containing JAK1 truncating mutations in the Cancer Cell Line Encyclopedia databank, 68% are gynecologic cancer cells. Within JAK1 the K142, P430, and K860 frame-shift mutations were identified as hot spot mutation sites. Sanger sequencing of cancer cell lines, primary tumors, and matched normal tissues confirmed the JAK1 mutations and showed that these mutations are somatic. JAK1 mediates interferon (IFN)-γ-regulated tumor immune surveillance. Functional assays show that JAK1 deficient cancer cells are defective in IFN-γ-induced LMP2 and TAP1 expression, loss of which inhibits presentation of tumor antigens. These findings identify recurrent JAK1 truncating mutations that could contribute to tumor immune evasion in gynecologic cancers, especially in endometrial cancer.
Figures
Similar articles
-
The mutation in the ATP-binding region of JAK1, identified in human uterine leiomyosarcomas, results in defective interferon-gamma inducibility of TAP1 and LMP2.Oncogene. 2006 Jul 6;25(29):4016-26. doi: 10.1038/sj.onc.1209434. Epub 2006 Feb 13. Oncogene. 2006. PMID: 16474838
-
Loss of interferon-gamma inducibility of TAP1 and LMP2 in a renal cell carcinoma cell line.Cancer Res. 2000 Oct 15;60(20):5789-96. Cancer Res. 2000. PMID: 11059775
-
Interferon induction of TAP1: the phosphatase SHP-1 regulates crossover between the IFN-alpha/beta and the IFN-gamma signal-transduction pathways.Circ Res. 1998 Oct 19;83(8):815-23. doi: 10.1161/01.res.83.8.815. Circ Res. 1998. PMID: 9776728
-
Mutations leading to constitutive active gp130/JAK1/STAT3 pathway.Cytokine Growth Factor Rev. 2015 Oct;26(5):499-506. doi: 10.1016/j.cytogfr.2015.07.010. Epub 2015 Jul 3. Cytokine Growth Factor Rev. 2015. PMID: 26188635 Review.
-
How do endogenous proteins become peptides and reach the endoplasmic reticulum.Curr Top Microbiol Immunol. 1998;232:15-47. doi: 10.1007/978-3-642-72045-1_2. Curr Top Microbiol Immunol. 1998. PMID: 9557392 Review. No abstract available.
Cited by
-
JAK1: Number one in the family; number one in inflammation?Rheumatology (Oxford). 2021 May 5;60(Suppl 2):ii3-ii10. doi: 10.1093/rheumatology/keab024. Rheumatology (Oxford). 2021. PMID: 33950229 Free PMC article.
-
Biallelic JAK1 mutations in immunodeficient patient with mycobacterial infection.Nat Commun. 2016 Dec 23;7:13992. doi: 10.1038/ncomms13992. Nat Commun. 2016. PMID: 28008925 Free PMC article.
-
Gynecological Cancers Caused by Deficient Mismatch Repair and Microsatellite Instability.Cancers (Basel). 2020 Nov 10;12(11):3319. doi: 10.3390/cancers12113319. Cancers (Basel). 2020. PMID: 33182707 Free PMC article. Review.
-
Acquired IFNγ resistance impairs anti-tumor immunity and gives rise to T-cell-resistant melanoma lesions.Nat Commun. 2017 May 31;8:15440. doi: 10.1038/ncomms15440. Nat Commun. 2017. PMID: 28561041 Free PMC article.
-
RETooling the RET Inhibitor Pralsetinib for ESR1 Fusion-Positive Breast Cancer and Beyond.Cancer Res. 2023 Oct 2;83(19):3159-3161. doi: 10.1158/0008-5472.CAN-23-1021. Cancer Res. 2023. PMID: 37779428 Free PMC article.
References
-
- Pao W. & Girard N. New driver mutations in non-small-cell lung cancer. Lancet Oncol 12, 175–80 (2011). - PubMed
-
- Gilliland D. G., Jordan C. T. & Felix C. A. The molecular basis of leukemia. Hematology (Am Soc Hematol Educ Program), 80–97 (2004). - PubMed
-
- Deininger M., Buchdunger E. & Druker B. J. The development of imatinib as a therapeutic agent for chronic myeloid leukemia. Blood 105, 2640–53 (2005). - PubMed
-
- Tefferi A. & Gilliland D. G. Oncogenes in myeloproliferative disorders. Cell Cycle 6, 550–66 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
