

doi: 10.1128/JVI.02608-13.
Epub 2013 Oct 23.
Guiding the immune response against influenza virus hemagglutinin toward the conserved stalk domain by hyperglycosylation of the globular head domain
Affiliations
- PMID: 24155380
- PMCID: PMC3911724
- DOI: 10.1128/JVI.02608-13
Item in Clipboard
Guiding the immune response against influenza virus hemagglutinin toward the conserved stalk domain by hyperglycosylation of the globular head domain
J Virol.
2014 Jan.
Abstract
Influenza virus hemagglutinin consists of a highly variable and immunodominant head domain and a more conserved but immunosubdominant stalk domain. We introduced seven N-linked glycosylation sites in the hemagglutinin head domain to shield the immunodominant antigenic sites. The hyperglycosylated hemagglutinin enhanced stalk-directed seroreactivity while dampening the head response in immunized mice. Upon influenza virus challenge, mice vaccinated with the hyperglycosylated hemagglutinin were better protected against morbidity and mortality than mice receiving the wild-type hemagglutinin.
Figures
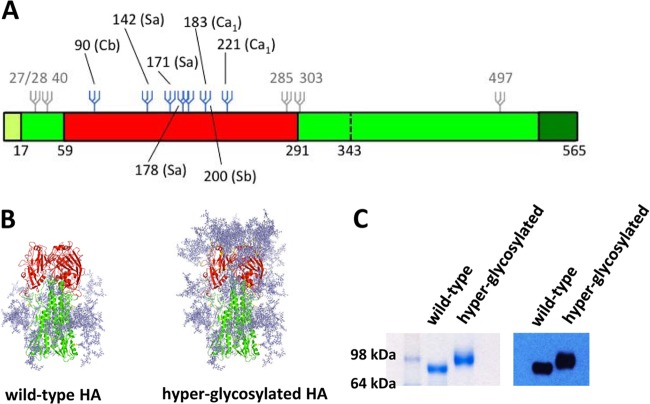
Covering antigenic sites of HA with N-linked glycosylation. (A) Schematic of the influenza HA and the position of introduced glycans. The signal peptide is shown in light green, the head and stalk domain of the ectodomain in red and green, respectively, and the transmembrane and cytoplasmic domains are depicted in dark green. Glycans indicated in gray are present in the wild-type HA. Additional glycans in the hyperglycosylated HA are depicted in blue. (B) Modeling of introduced glycans on the crystal structure of the PR/8 HA (PDB 1RU7). Original and additional glycans were modeled on the crystal structure of the PR/8 HA using GlyProt (37) and drawn using PyMol (DeLano Scientific). Because of modeling limitations, only 6 out of the 7 added potential N-linked glycans are present on the hyperglycosylated HA model. (C) The ectodomain of the wild-type HA and the hyperglycosylated HA containing the T4-foldon trimerization domain and a His tag were expressed in 293T cells, and purified glycoproteins were run on an SDS-PAGE polyacrylamide gel under denaturing and reducing conditions. A clear increase in molecular weight is present for the hyperglycosylated HA compared to the wild-type HA, indicating the presence of additional glycosylation.
Hyperglycosylated HA expresses well at the surface of transfected mammalian cells and is able to bind stalk antibodies. Plasmids encoding wild-type or hyperglycosylated full-length HA were transfected into 293T cells. The HA was stained 24 h posttransfection with PR8 antiserum or stalk-directed antibodies KB2 (28), C179 (29), 6F12 (11), and CR6261 (7) (A) or head-directed antibodies PY102 (38), 17, 43, 45, and 2 (B). Monoclonal antibodies 17, 43, 45, and 2 were generated for this research as described before (11). The indicated binding epitope at the PR/8 HA was mapped using escape studies in eggs for each newly generated HA head antibody as described previously (4). Binding of stalk-directed antibodies to the hyperglycosylated HA is not changed compared to that of the wild-type HA. In contrast, introduction of glycosylation sites in the antigenic sites of the globular head domain of the HA abrogated binding of head-directed antibodies. (C) Binding of five different head-directed antibodies to purified, soluble HA was tested in ELISAs. Head-directed antibodies did not bind to hyperglycosylated HA, while binding of stalk-directed antibody KB2 was not affected.
Immunization of mice with the hyperglycosylated HA induces high titers of stalk-directed antibodies against distinct seasonal H1N1 and pandemic H5N1 viruses. Sera originating from mice vaccinated with the wild-type or hyperglycosylated HAs were assayed by ELISA for reactivity against purified viruses: A/Puerto Rico/8/34, cH5/1, A/USSR/92/77, A/Brisbane/59/07, and A/Vietnam/1203/04. Sera originating from mice vaccinated with the hyperglycosylated HA were less reactive to virus containing the full-length PR/8 HA compared to mice vaccinated with the wild-type HA after two (A) and three (B) immunizations. In contrast, seroreactivity to a virus bearing the cH5/1 HA, which consists of a subtype H5 head domain and the PR/8 stalk domain, was higher for sera originating from mice vaccinated with the hyperglycosylated HA than in mice vaccinated with the wild-type HA. This difference in reactivity was observed after two immunizations (C) and became more apparent after three immunizations (D). The seroreactivity to distinct H1N1 viruses (E, F) and an H5N1 virus (G) was also enhanced in mice vaccinated with the hyperglycosylated HA compared to that in mice immunized with the wild-type HA.
Mice immunized with the hyperglycosylated HA show protection against morbidity and mortality upon challenge with a cH9/1 virus. (A) Mice (n = 5) were challenged with 20 mLD50 of a virus containing the cH9/1 HA and weighed daily. Vaccination with the hyperglycosylated HA protected mice better from morbidity as measured by weight loss (A) and mortality (B) compared to mice vaccinated with the wild-type HA. Significant differences between the hyperglycosylated HA and the wild-type HA-vaccinated mice were observed at the indicated points. Statistical significance for weight curves was calculated using multiple unpaired t tests correcting for multiple comparisons with the Holm-Sidak method (*, P < 0.001; Prism version 6.0b; GraphPad Software). Statistical significance for the Kaplan-Meier survival curve was calculated with the Mantel-Cox test (*, P < 0.05; Prism version 6.0b; GraphPad Software).
References
-
- Margine I, Hai R, Albrecht RA, Obermoser G, Harrod AC, Banchereau J, Palucka K, Garcia-Sastre A, Palese P, Treanor JJ, Krammer F. 2013. H3N2 influenza virus infection induces broadly reactive hemagglutinin stalk antibodies in humans and mice. J. Virol. 87:4728–4737. 10.1128/JVI.03509-12 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
