

Genomewide mapping and screening of Kaposi's sarcoma-associated herpesvirus (KSHV) 3' untranslated regions identify bicistronic and polycistronic viral transcripts as frequent targets of KSHV microRNAs
- PMID: 24155407
- PMCID: PMC3911756
- DOI: 10.1128/JVI.02689-13
Genomewide mapping and screening of Kaposi's sarcoma-associated herpesvirus (KSHV) 3' untranslated regions identify bicistronic and polycistronic viral transcripts as frequent targets of KSHV microRNAs
Abstract
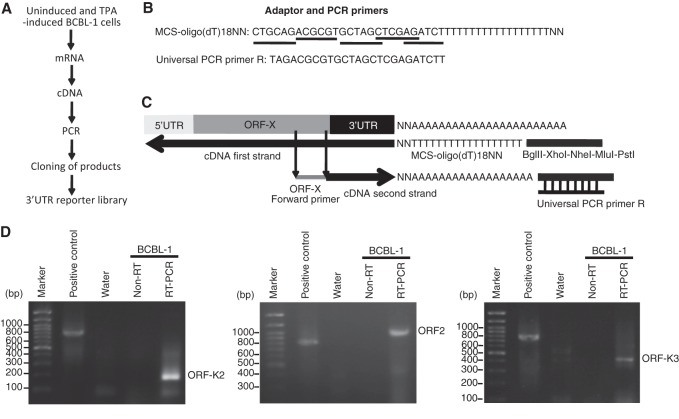
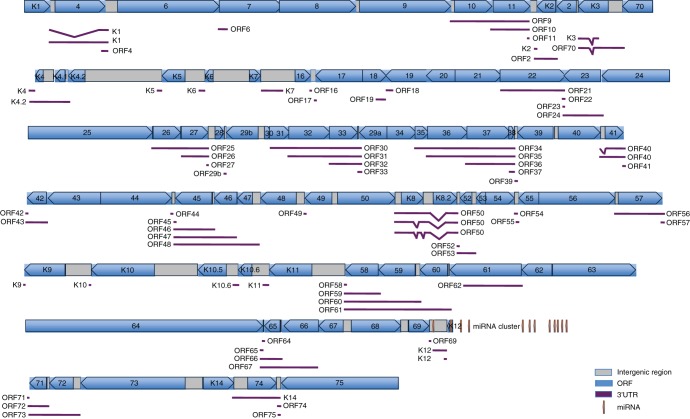
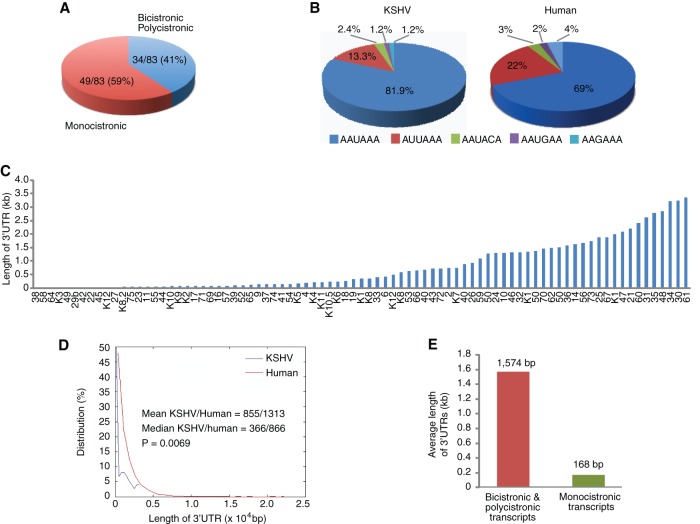
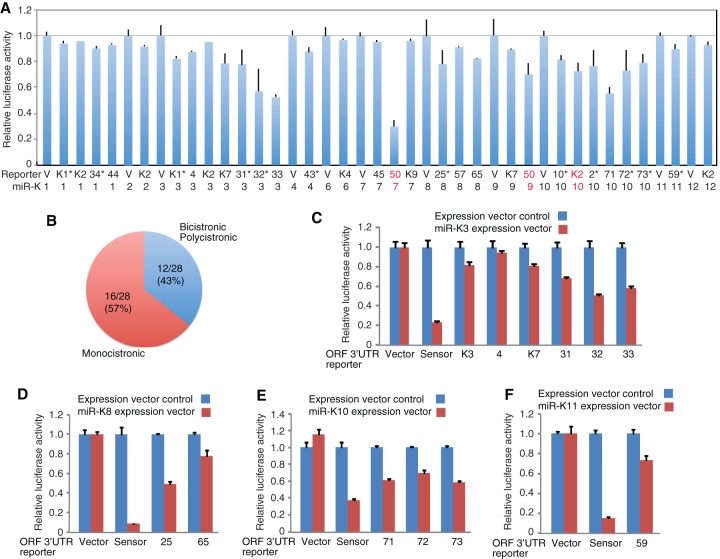
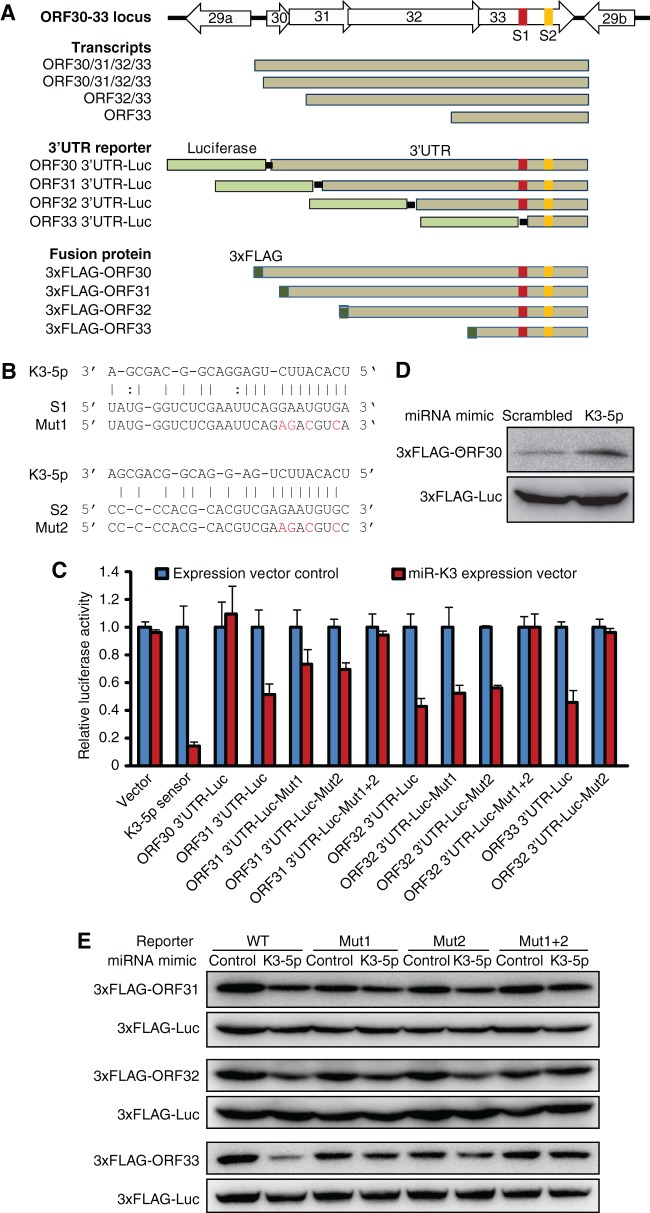
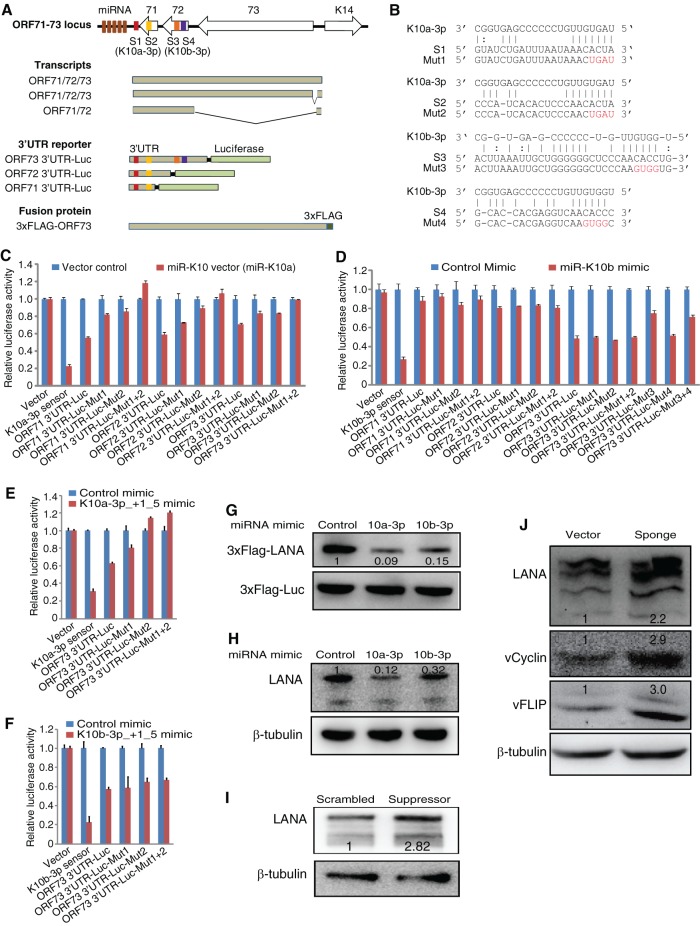
Kaposi's sarcoma-associated herpesvirus (KSHV) encodes over 90 genes and 25 microRNAs (miRNAs). The KSHV life cycle is tightly regulated to ensure persistent infection in the host. In particular, miRNAs, which primarily exert their effects by binding to the 3' untranslated regions (3'UTRs) of target transcripts, have recently emerged as key regulators of KSHV life cycle. Although studies with RNA cross-linking immunoprecipitation approach have identified numerous targets of KSHV miRNAs, few of these targets are of viral origin because most KSHV 3'UTRs have not been characterized. Thus, the extents of viral genes targeted by KSHV miRNAs remain elusive. Here, we report the mapping of the 3'UTRs of 74 KSHV genes and the effects of KSHV miRNAs on the control of these 3'UTR-mediated gene expressions. This analysis reveals new bicistronic and polycistronic transcripts of KSHV genes. Due to the 5'-distal open reading frames (ORFs), KSHV bicistronic or polycistronic transcripts have significantly longer 3'UTRs than do KSHV monocistronic transcripts. Furthermore, screening of the 3'UTR reporters has identified 28 potential new targets of KSHV miRNAs, of which 11 (39%) are bicistronic or polycistronic transcripts. Reporter mutagenesis demonstrates that miR-K3 specifically targets ORF31-33 transcripts at the lytic locus via two binding sites in the ORF33 coding region, whereas miR-K10a-3p and miR-K10b-3p and their variants target ORF71-73 transcripts at the latent locus through distinct binding sites in both 5'-distal ORFs and intergenic regions. Our results indicate that KSHV miRNAs frequently target the 5'-distal coding regions of bicistronic or polycistronic transcripts and highlight the unique features of KSHV miRNAs in regulating gene expression and life cycle.
Figures
Similar articles
-
Viral miRNA targeting of bicistronic and polycistronic transcripts.Curr Opin Virol. 2014 Aug;7:66-72. doi: 10.1016/j.coviro.2014.04.004. Epub 2014 May 10. Curr Opin Virol. 2014. PMID: 24821460 Free PMC article. Review.
-
Modified Cross-Linking, Ligation, and Sequencing of Hybrids (qCLASH) Identifies Kaposi's Sarcoma-Associated Herpesvirus MicroRNA Targets in Endothelial Cells.J Virol. 2018 Mar 28;92(8):e02138-17. doi: 10.1128/JVI.02138-17. Print 2018 Apr 15. J Virol. 2018. PMID: 29386283 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus microRNAs repress breakpoint cluster region protein expression, enhance Rac1 activity, and increase in vitro angiogenesis.J Virol. 2015 Apr;89(8):4249-61. doi: 10.1128/JVI.03687-14. Epub 2015 Jan 28. J Virol. 2015. PMID: 25631082 Free PMC article.
-
Kaposi's Sarcoma-Associated Herpesvirus MicroRNAs Target GADD45B To Protect Infected Cells from Cell Cycle Arrest and Apoptosis.J Virol. 2017 Jan 18;91(3):e02045-16. doi: 10.1128/JVI.02045-16. Print 2017 Feb 1. J Virol. 2017. PMID: 27852859 Free PMC article.
-
KSHV microRNAs: Tricks of the Devil.Trends Microbiol. 2017 Aug;25(8):648-661. doi: 10.1016/j.tim.2017.02.002. Epub 2017 Mar 2. Trends Microbiol. 2017. PMID: 28259385 Free PMC article. Review.
Cited by
-
Epstein-Barr Virus miR-BHRF1-3 Targets the BZLF1 3'UTR and Regulates the Lytic Cycle.J Virol. 2022 Feb 23;96(4):e0149521. doi: 10.1128/JVI.01495-21. Epub 2021 Dec 8. J Virol. 2022. PMID: 34878852 Free PMC article.
-
Inhibition of Kaposi's sarcoma-associated herpesvirus lytic replication by HIV-1 Nef and cellular microRNA hsa-miR-1258.J Virol. 2014 May;88(9):4987-5000. doi: 10.1128/JVI.00025-14. Epub 2014 Feb 19. J Virol. 2014. PMID: 24554664 Free PMC article.
-
The Marek's Disease Virus Unique Gene MDV082 Is Dispensable for Virus Replication but Contributes to a Rapid Disease Onset.J Virol. 2021 Jul 12;95(15):e0013121. doi: 10.1128/JVI.00131-21. Epub 2021 Jul 12. J Virol. 2021. PMID: 34011541 Free PMC article.
-
Expression of the Antisense-to-Latency Transcript Long Noncoding RNA in Kaposi's Sarcoma-Associated Herpesvirus.J Virol. 2017 Jan 31;91(4):e01698-16. doi: 10.1128/JVI.01698-16. Print 2017 Feb 15. J Virol. 2017. PMID: 27928018 Free PMC article.
-
HIV-1 Nef and KSHV oncogene K1 synergistically promote angiogenesis by inducing cellular miR-718 to regulate the PTEN/AKT/mTOR signaling pathway.Nucleic Acids Res. 2014 Sep;42(15):9862-79. doi: 10.1093/nar/gku583. Epub 2014 Aug 7. Nucleic Acids Res. 2014. PMID: 25104021 Free PMC article.
References
-
- Pi H, Lee LW, Lo SJ. 2009. New insights into polycistronic transcripts in eukaryotes. Chang Gung Med. J. 32:494–498 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- CA132637/CA/NCI NIH HHS/United States
- R01 CA096512/CA/NCI NIH HHS/United States
- R01 CA124332/CA/NCI NIH HHS/United States
- AI073099/AI/NIAID NIH HHS/United States
- CA31363/CA/NCI NIH HHS/United States
- CA096512/CA/NCI NIH HHS/United States
- P01 CA180779/CA/NCI NIH HHS/United States
- R01 AI073099/AI/NIAID NIH HHS/United States
- R01 CA031363/CA/NCI NIH HHS/United States
- CA180779/CA/NCI NIH HHS/United States
- R21 AI105909/AI/NIAID NIH HHS/United States
- R01 CA082057/CA/NCI NIH HHS/United States
- R01 CA115284/CA/NCI NIH HHS/United States
- R01 CA177377/CA/NCI NIH HHS/United States
- CA082057/CA/NCI NIH HHS/United States
- CA115284/CA/NCI NIH HHS/United States
- AI105809/AI/NIAID NIH HHS/United States
- R01 CA132637/CA/NCI NIH HHS/United States
- CA124332/CA/NCI NIH HHS/United States
- CA177377/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
