

Activation of high and low affinity dopamine receptors generates a closed loop that maintains a conductance ratio and its activity correlate
- PMID: 24155696
- PMCID: PMC3805135
- DOI: 10.3389/fncir.2013.00169
Activation of high and low affinity dopamine receptors generates a closed loop that maintains a conductance ratio and its activity correlate
Abstract
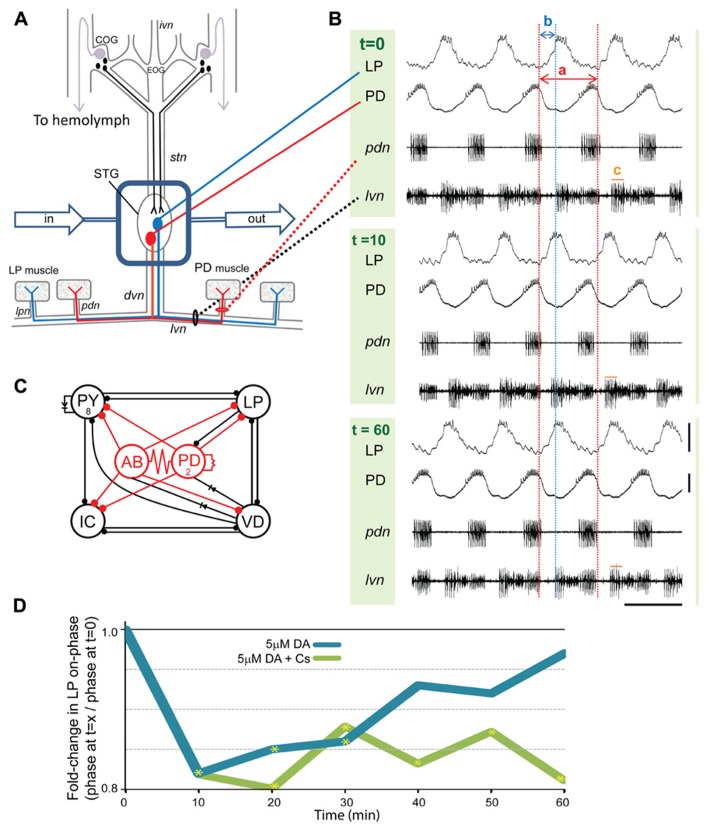
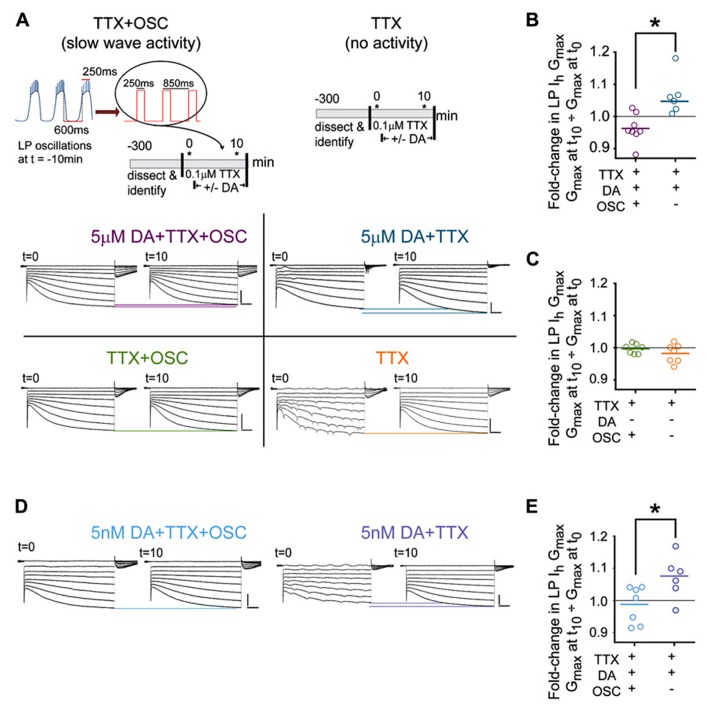
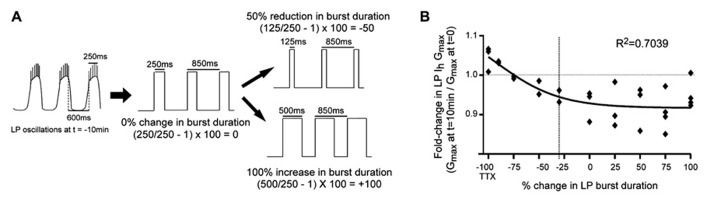
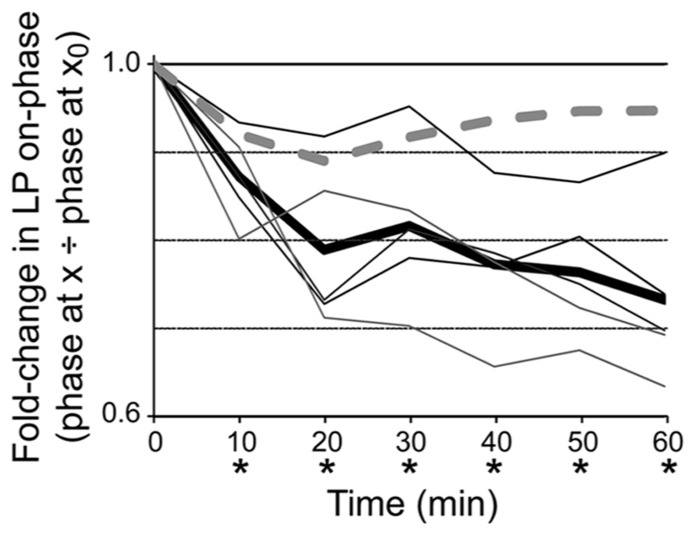
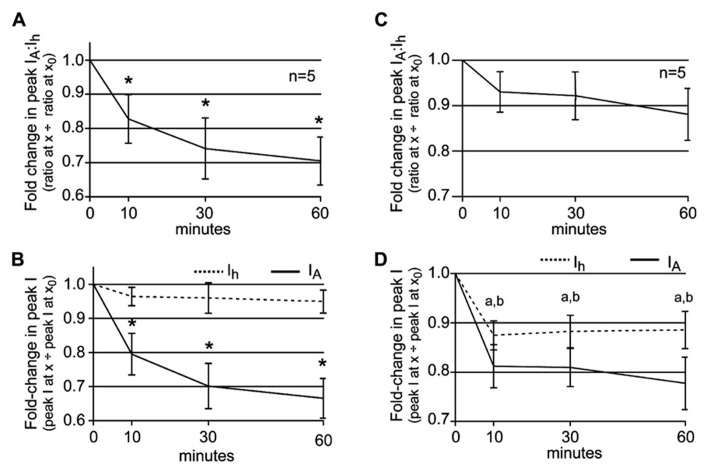
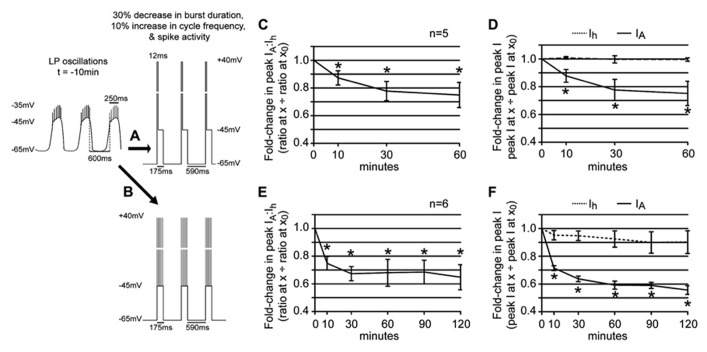
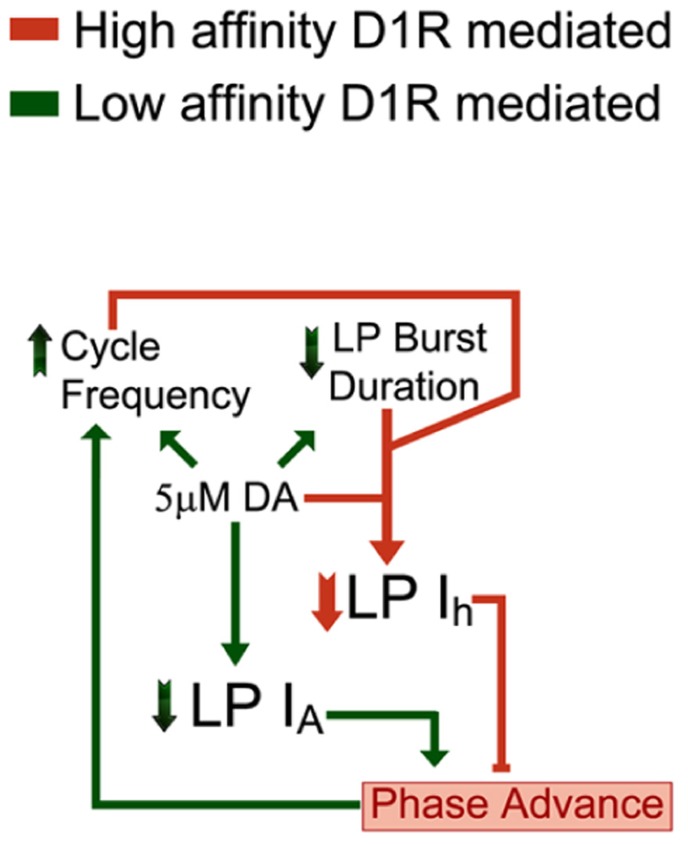
Neuromodulators alter network output and have the potential to destabilize a circuit. The mechanisms maintaining stability in the face of neuromodulation are not well described. Using the pyloric network in the crustacean stomatogastric nervous system, we show that dopamine (DA) does not simply alter circuit output, but activates a closed loop in which DA-induced alterations in circuit output consequently drive a change in an ionic conductance to preserve a conductance ratio and its activity correlate. DA acted at low affinity type 1 receptors (D1Rs) to induce an immediate modulatory decrease in the transient potassium current (IA) of a pyloric neuron. This, in turn, advanced the activity phase of that component neuron, which disrupted its network function and thereby destabilized the circuit. DA simultaneously acted at high affinity D1Rs on the same neuron to confer activity-dependence upon the hyperpolarization activated current (Ih) such that the DA-induced changes in activity subsequently reduced Ih. This DA-enabled, activity-dependent, intrinsic plasticity exactly compensated for the modulatory decrease in IA to restore the IA:Ih ratio and neuronal activity phase, thereby closing an open loop created by the modulator. Activation of closed loops to preserve conductance ratios may represent a fundamental operating principle neuromodulatory systems use to ensure stability in their target networks.
Keywords: HCN channel; activity-dependent intrinsic plasticity; metamodulation; metaplasticity; pyloric network; stomatogastric.
Figures
References
-
- Baro D. J., Levini R. M., Kim M. T., Willms A. R., Lanning C. C., Rodriguez H. E., et al. (1997). Quantitative single-cell-reverse transcription-PCR demon-strates that A-current magnitude varies as a linear function of shal gene expression in identified stomatogastric neurons. J. Neurosci. 17 6597–6610 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources