

IL-21 signalling via STAT3 primes human naive B cells to respond to IL-2 to enhance their differentiation into plasmablasts
- PMID: 24159173
- PMCID: PMC3854113
- DOI: 10.1182/blood-2013-06-506865
IL-21 signalling via STAT3 primes human naive B cells to respond to IL-2 to enhance their differentiation into plasmablasts
Abstract
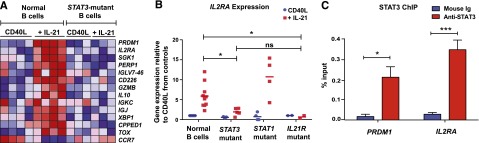
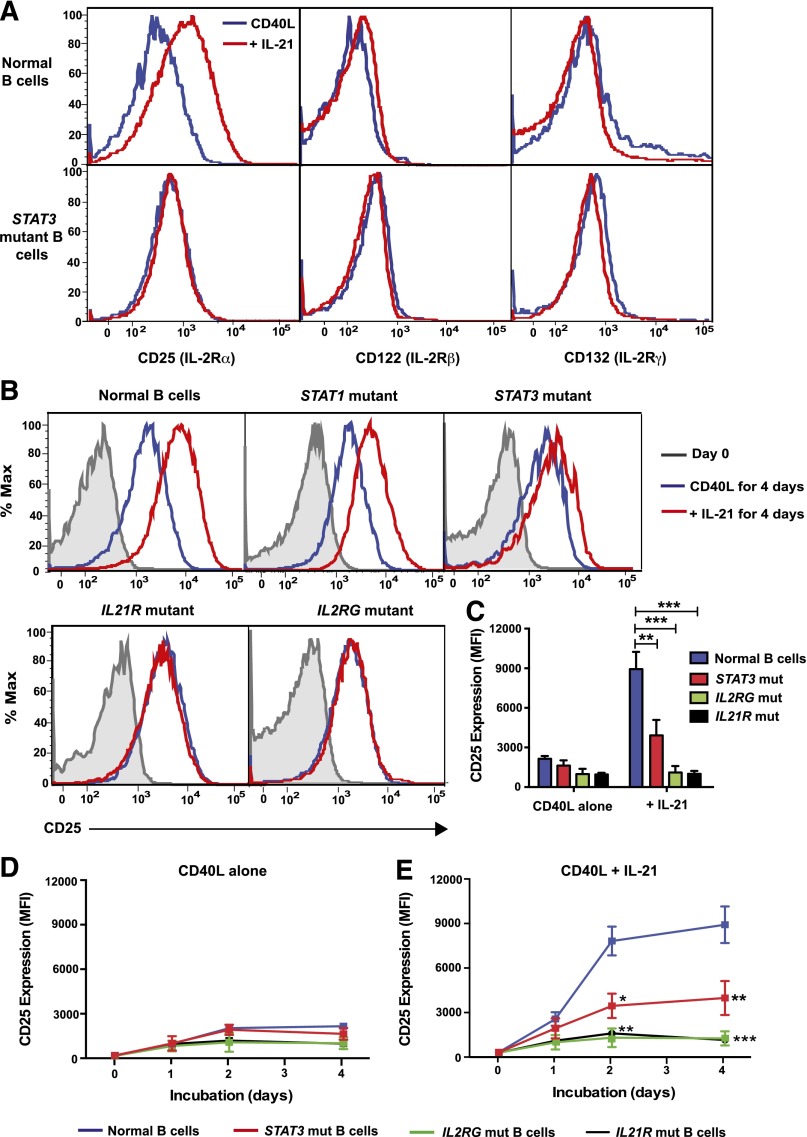
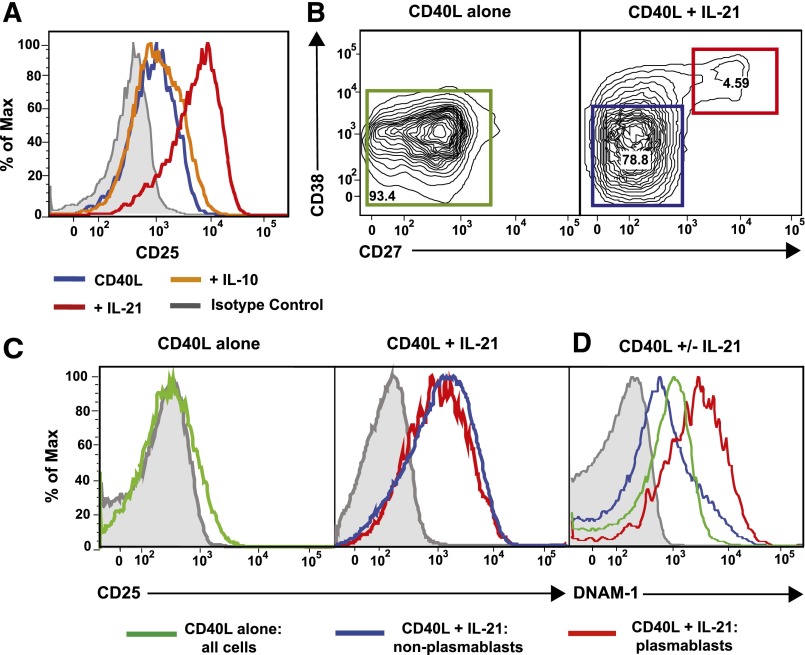
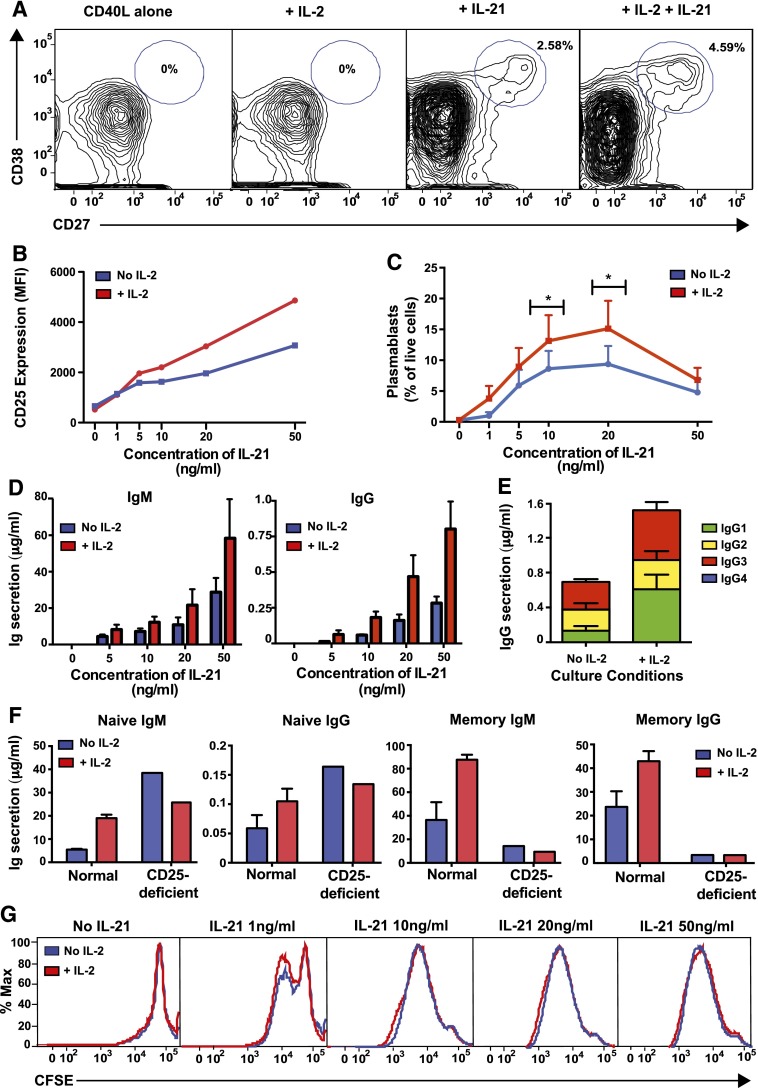
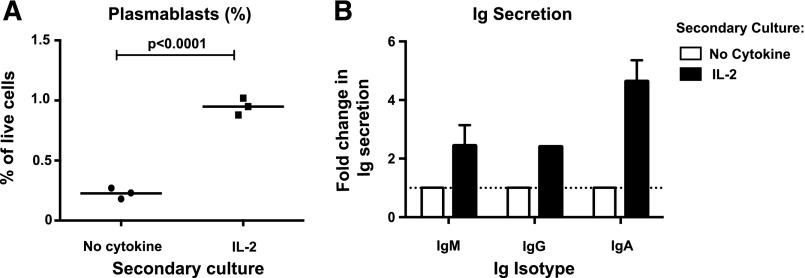
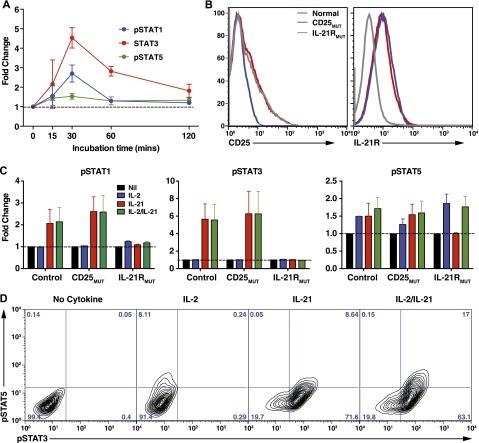
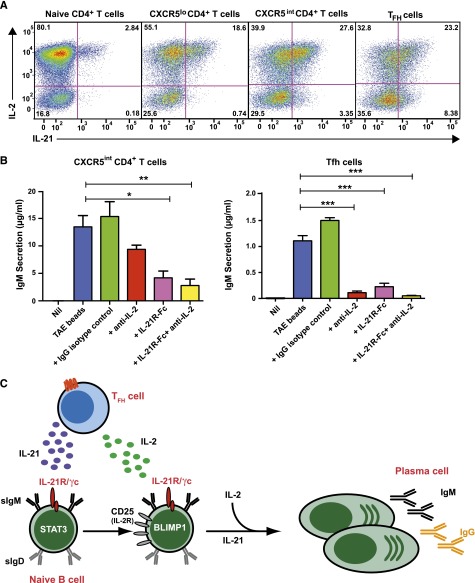
B-cell responses are guided by the integration of signals through the B-cell receptor (BCR), CD40, and cytokine receptors. The common γ chain (γc)-binding cytokine interleukin (IL)-21 drives humoral immune responses via STAT3-dependent induction of transcription factors required for plasma cell generation. We investigated additional mechanisms by which IL-21/STAT3 signaling modulates human B-cell responses by studying patients with STAT3 mutations. IL-21 strongly induced CD25 (IL-2Rα) in normal, but not STAT3-deficient, CD40L-stimulated naïve B cells. Chromatin immunoprecipitation confirmed IL2RA as a direct target of STAT3. IL-21-induced CD25 expression was also impaired on B cells from patients with IL2RG or IL21R mutations, confirming a requirement for intact IL-21R signaling in this process. IL-2 increased plasmablast generation and immunoglobulin secretion from normal, but not CD25-deficient, naïve B cells stimulated with CD40L/IL-21. IL-2 and IL-21 were produced by T follicular helper cells, and neutralizing both cytokines abolished the B-cell helper capacity of these cells. Our results demonstrate that IL-21, via STAT3, sensitizes B cells to the stimulatory effects of IL-2. Thus, IL-2 may play an adjunctive role in IL-21-induced B-cell differentiation. Lack of this secondary effect of IL-21 may amplify the humoral immunodeficiency in patients with mutations in STAT3, IL2RG, or IL21R due to impaired responsiveness to IL-21.
Figures
References
-
- Tangye SG, Tarlinton DM. Memory B cells: effectors of long-lived immune responses. Eur J Immunol. 2009;39(8):2065–2075. - PubMed
-
- Goodnow CC, Vinuesa CG, Randall KL, Mackay F, Brink R. Control systems and decision making for antibody production. Nat Immunol. 2010;11(8):681–688. - PubMed
-
- Victora GD, Nussenzweig MC. Germinal centers. Annu Rev Immunol. 2012;30:429–457. - PubMed
-
- Crotty S. Follicular helper CD4 T cells (TFH). Annu Rev Immunol. 2011;29:621–663. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
