

Wnt secretion is required to maintain high levels of Wnt activity in colon cancer cells
- PMID: 24162018
- PMCID: PMC3826636
- DOI: 10.1038/ncomms3610
Wnt secretion is required to maintain high levels of Wnt activity in colon cancer cells
Abstract
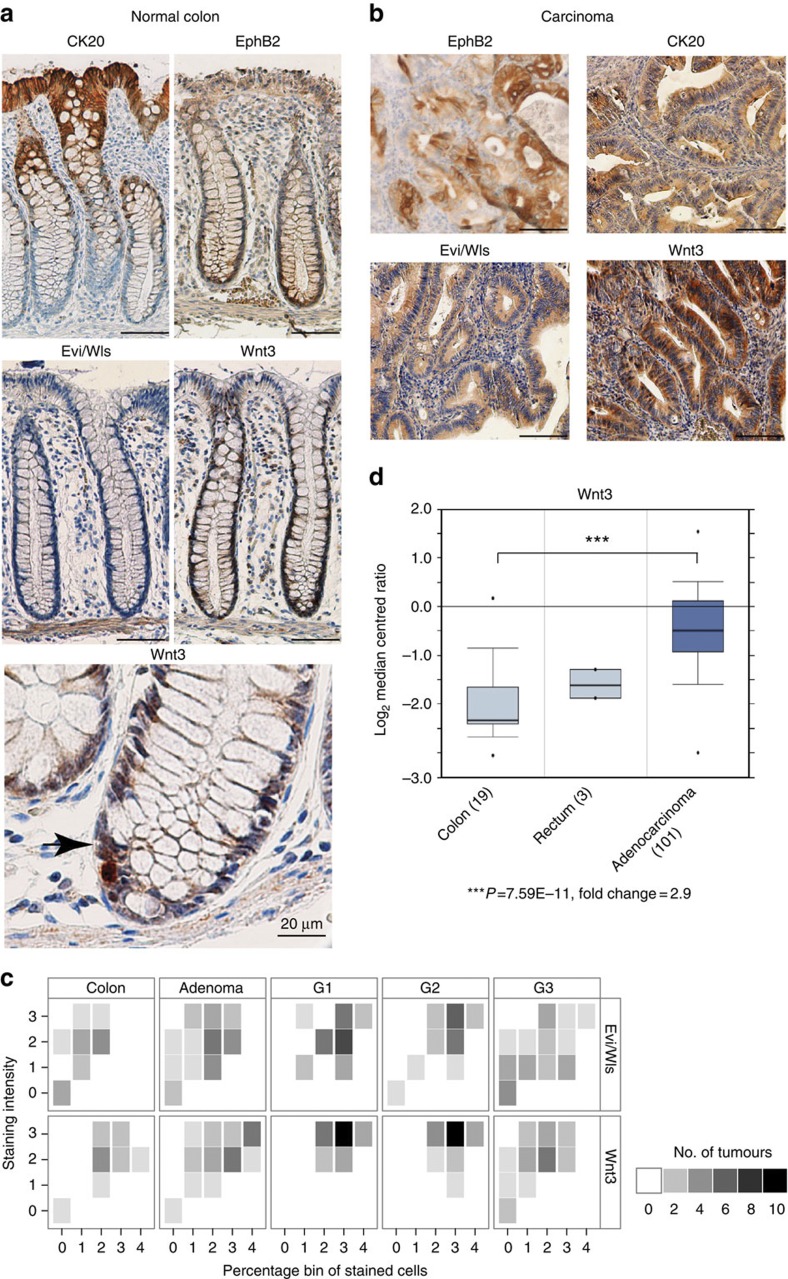
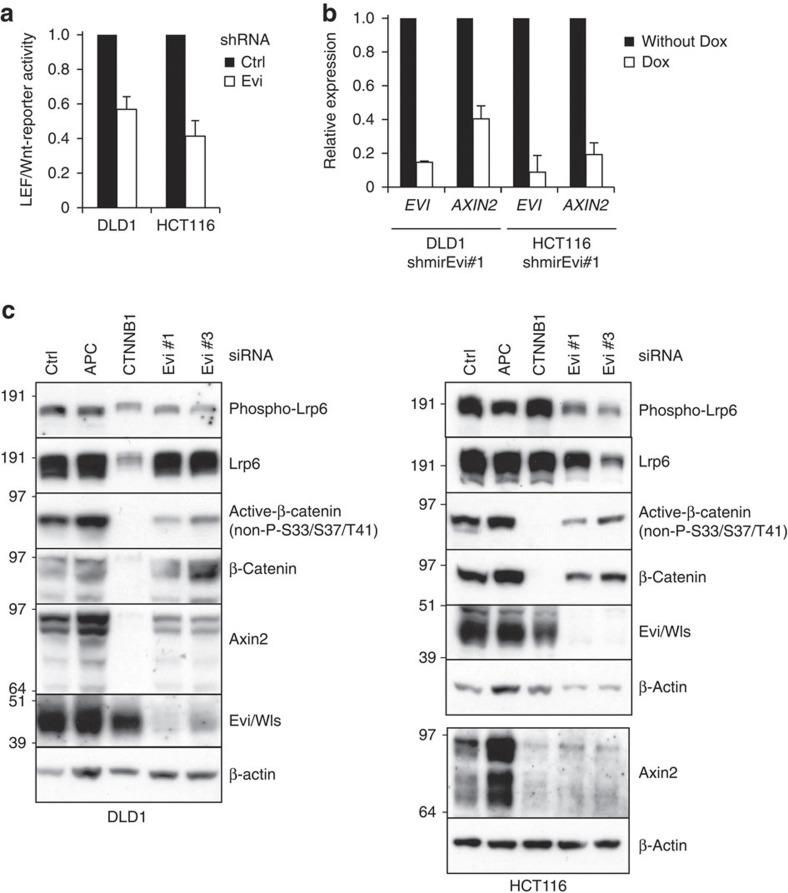
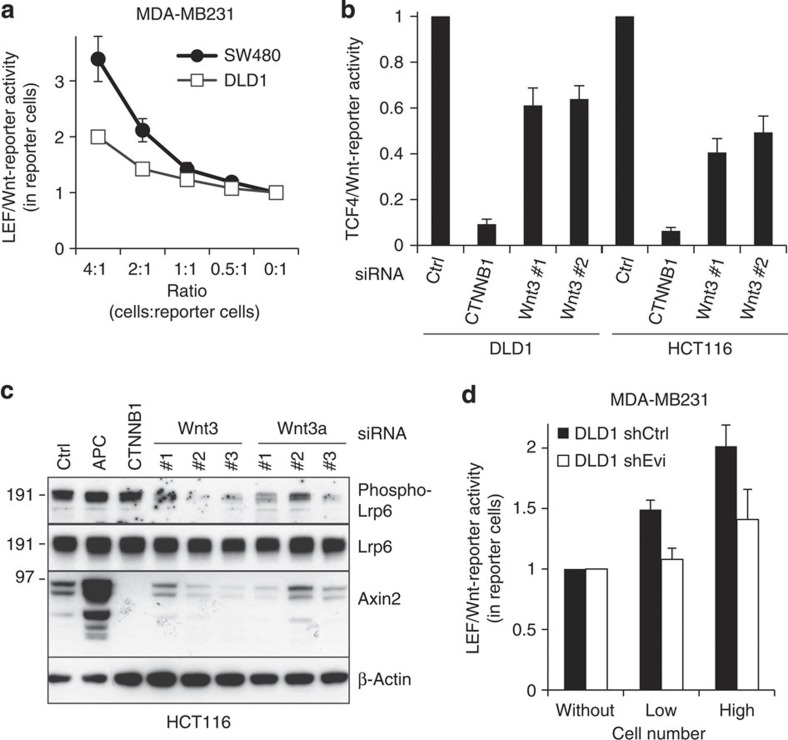
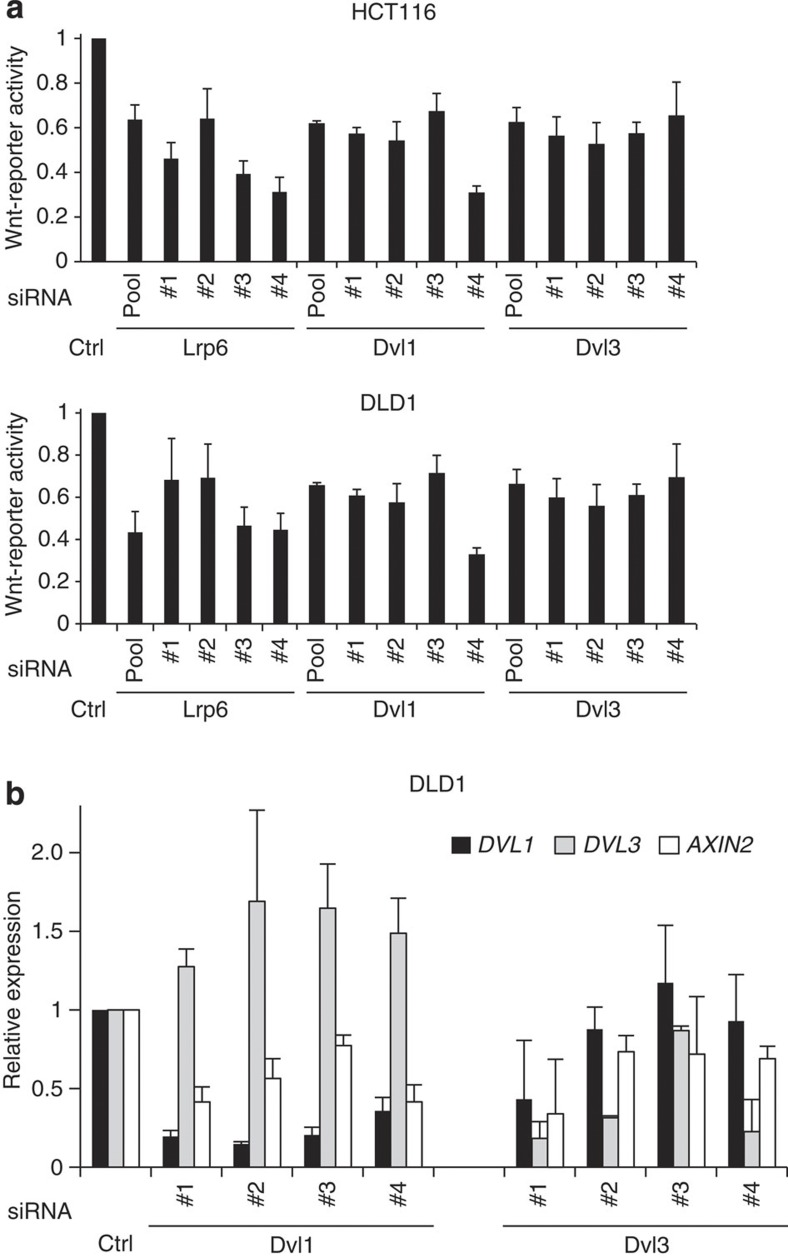
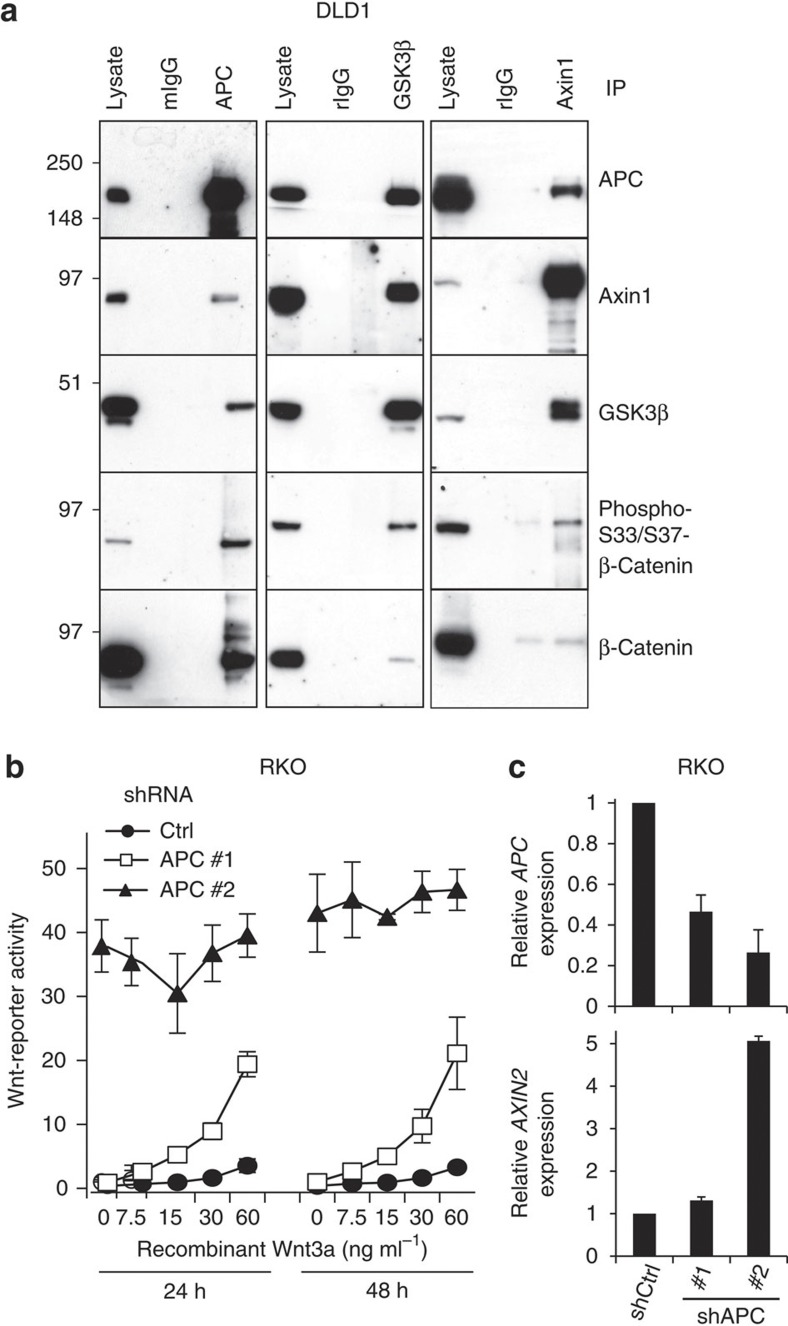
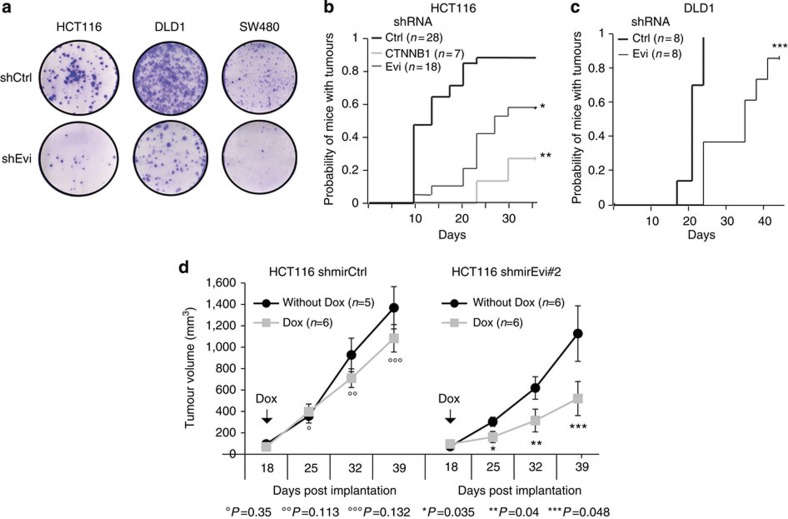
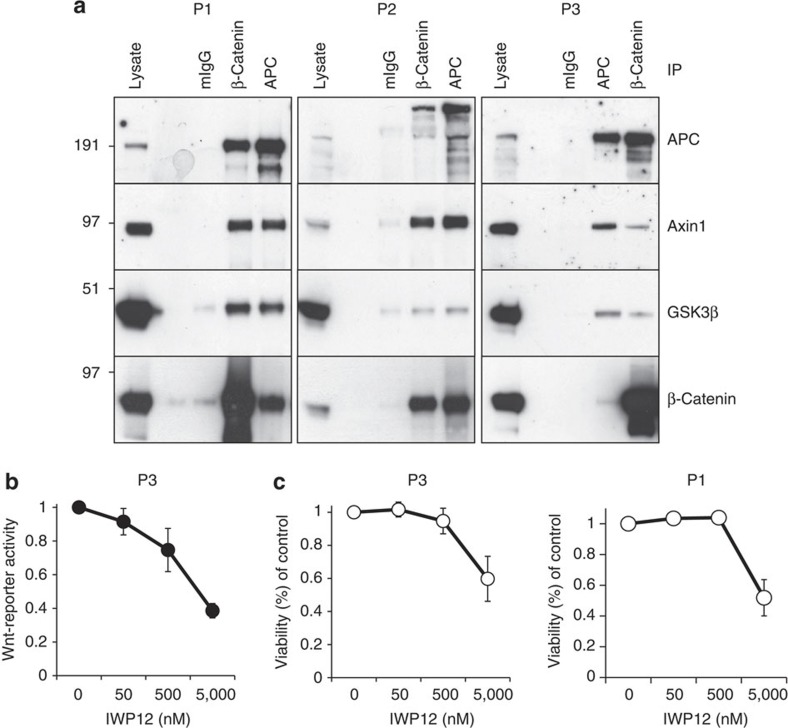
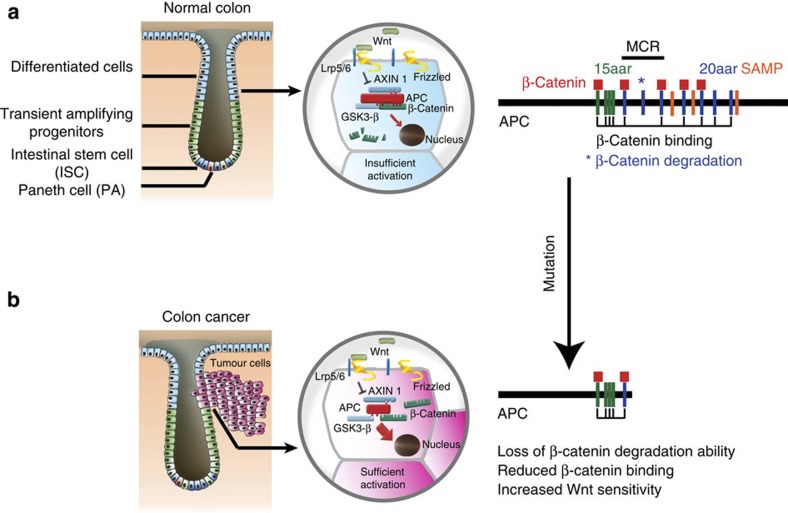
Aberrant regulation of the Wnt/β-catenin pathway has an important role during the onset and progression of colorectal cancer, with over 90% of cases of sporadic colon cancer featuring mutations in APC or β-catenin. However, it has remained a point of controversy whether these mutations are sufficient to activate the pathway or require additional upstream signals. Here we show that colorectal tumours express elevated levels of Wnt3 and Evi/Wls/GPR177. We found that in colon cancer cells, even in the presence of mutations in APC or β-catenin, downstream signalling remains responsive to Wnt ligands and receptor proximal signalling. Furthermore, we demonstrate that truncated APC proteins bind β-catenin and key components of the destruction complex. These results indicate that cells with mutations in APC or β-catenin depend on Wnt ligands and their secretion for a sufficient level of β-catenin signalling, which potentially opens new avenues for therapeutic interventions by targeting Wnt secretion via Evi/Wls.
Figures

Similar articles
-
Vitamin D receptor deficiency enhances Wnt/β-catenin signaling and tumor burden in colon cancer.PLoS One. 2011;6(8):e23524. doi: 10.1371/journal.pone.0023524. Epub 2011 Aug 15. PLoS One. 2011. PMID: 21858154 Free PMC article.
-
The Wnt/β-catenin/T-cell factor 4 pathway up-regulates high-mobility group A1 expression in colon cancer.Cell Biochem Funct. 2013 Apr;31(3):228-36. doi: 10.1002/cbf.2876. Epub 2012 Sep 7. Cell Biochem Funct. 2013. PMID: 22961697 Free PMC article.
-
The COP9 signalosome mediates beta-catenin degradation by deneddylation and blocks adenomatous polyposis coli destruction via USP15.J Mol Biol. 2009 Aug 28;391(4):691-702. doi: 10.1016/j.jmb.2009.06.066. Epub 2009 Jul 1. J Mol Biol. 2009. PMID: 19576224
-
Wnt signaling and colon tumorigenesis--a view from the periphery.Exp Cell Res. 2011 Nov 15;317(19):2748-58. doi: 10.1016/j.yexcr.2011.08.010. Epub 2011 Aug 22. Exp Cell Res. 2011. PMID: 21884696 Review.
-
Cytoskeleton out of the cupboard: colon cancer and cytoskeletal changes induced by loss of APC.Nat Rev Cancer. 2006 Dec;6(12):967-74. doi: 10.1038/nrc2010. Epub 2006 Nov 9. Nat Rev Cancer. 2006. PMID: 17093505 Review.
Cited by
-
Inhibition of protein tyrosine phosphatase receptor type F suppresses Wnt signaling in colorectal cancer.Oncogene. 2020 Oct;39(44):6789-6801. doi: 10.1038/s41388-020-01472-z. Epub 2020 Sep 24. Oncogene. 2020. PMID: 32973331 Free PMC article.
-
Differential expression of mucins in Middle Eastern patients with colorectal cancer.Oncol Lett. 2016 Jul;12(1):393-400. doi: 10.3892/ol.2016.4672. Epub 2016 Jun 1. Oncol Lett. 2016. PMID: 27347157 Free PMC article.
-
From Crypts to Cancer: A Holistic Perspective on Colorectal Carcinogenesis and Therapeutic Strategies.Int J Mol Sci. 2024 Aug 30;25(17):9463. doi: 10.3390/ijms25179463. Int J Mol Sci. 2024. PMID: 39273409 Free PMC article. Review.
-
Wnt ligand-dependent activation of the negative feedback regulator Nkd1.Mol Biol Cell. 2015 Jun 15;26(12):2375-84. doi: 10.1091/mbc.E14-12-1648. Epub 2015 Apr 22. Mol Biol Cell. 2015. PMID: 25904337 Free PMC article.
-
Spatial Patterning from an Integrated Wnt/β-catenin and Notch/Delta Gene Circuit.Annu Int Conf IEEE Eng Med Biol Soc. 2018 Jul;2018:5022-5025. doi: 10.1109/EMBC.2018.8513462. Annu Int Conf IEEE Eng Med Biol Soc. 2018. PMID: 30441469 Free PMC article.
References
-
- Logan C. Y. & Nusse R. The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 20, 781–810 (2004). - PubMed
-
- Morin P. J. et al. Activation of beta-catenin-Tcf signaling in colon cancer by mutations in beta-catenin or APC. Science 275, 1787–1790 (1997). - PubMed
-
- Reya T. & Clevers H. Wnt signalling in stem cells and cancer. Nature 434, 843–850 (2005). - PubMed
-
- Humphries A. & Wright N. A. Colonic crypt organization and tumorigenesis. Nat. Rev. Cancer 8, 415–424 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
