

Roles of intracellular cyclic AMP signal transduction in the capacitation and subsequent hyperactivation of mouse and boar spermatozoa
- PMID: 24162806
- PMCID: PMC3934125
- DOI: 10.1262/jrd.2013-056
Roles of intracellular cyclic AMP signal transduction in the capacitation and subsequent hyperactivation of mouse and boar spermatozoa
Abstract
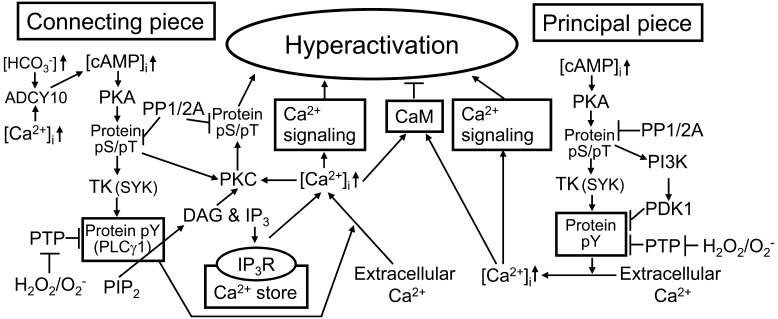
It is not until accomplishment of a variety of molecular changes during the transit through the female reproductive tract that mammalian spermatozoa are capable of exhibiting highly activated motility with asymmetric whiplash beating of the flagella (hyperactivation) and undergoing acrosomal exocytosis in the head (acrosome reaction). These molecular changes of the spermatozoa are collectively termed capacitation and promoted by bicarbonate, calcium and cholesterol acceptors. Such capacitation-promoting factors can stimulate intracellular cyclic AMP (cAMP) signal transduction in the spermatozoa. Meanwhile, hyperactivation and the acrosome reaction are essential to sperm fertilization with oocytes and are apparently triggered by a sufficient increase of intracellular Ca²⁺ in the sperm flagellum and head, respectively. Thus, it is necessary to investigate the relationship between cAMP signal transduction and calcium signaling cascades in the spermatozoa for the purpose of understanding the molecular basis of capacitation. In this review, I cover updated insights regarding intracellular cAMP signal transduction, the acrosome reaction and flagellar motility in mammalian spermatozoa and then account for possible roles of intracellular cAMP signal transduction in the capacitation and subsequent hyperactivation of mouse and boar spermatozoa.
Figures
References
-
- Bedford JM. Maturation, transport and fate of spermatozoa in the epididymis. In: Astwood EB, Greep RO (eds.), Handbook of Physiology, Section 7: Washington DC: American Physiological Society; 1975: 303–318.
-
- Orgebin-Crist MC, Danzo BJ, Davies J. Endocrine control of the development and maintenance of sperm fertilizing ability in the epididymis. In: Astwood EB, Greep RO (eds.), Handbook of Physiology, Section 7. Washington DC: American Physiological Society; 1975: 319–338.
-
- Hammerstedt RH, Parks JE. Changes in sperm surfaces associated with epididymal transit. J Reprod Fertil Suppl 1987; 34: 133–149 - PubMed
-
- Yanagimachi R. Mammalian fertilization. In: Knobil E, Neill JD (eds.), The Physiology of Reproduction. 2nd edition. New York: Raven Press; 1994: 189–317.
-
- Lewis B, Aitken RJ. Impact of epididymal maturation on the tyrosine phosphorylation patterns exhibited by rat spermatozoa. Biol Reprod 2001; 64: 1545–1556 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous