

An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens
- PMID: 24163310
- PMCID: PMC3877811
- DOI: 10.1105/tpc.113.117648
An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens
Abstract
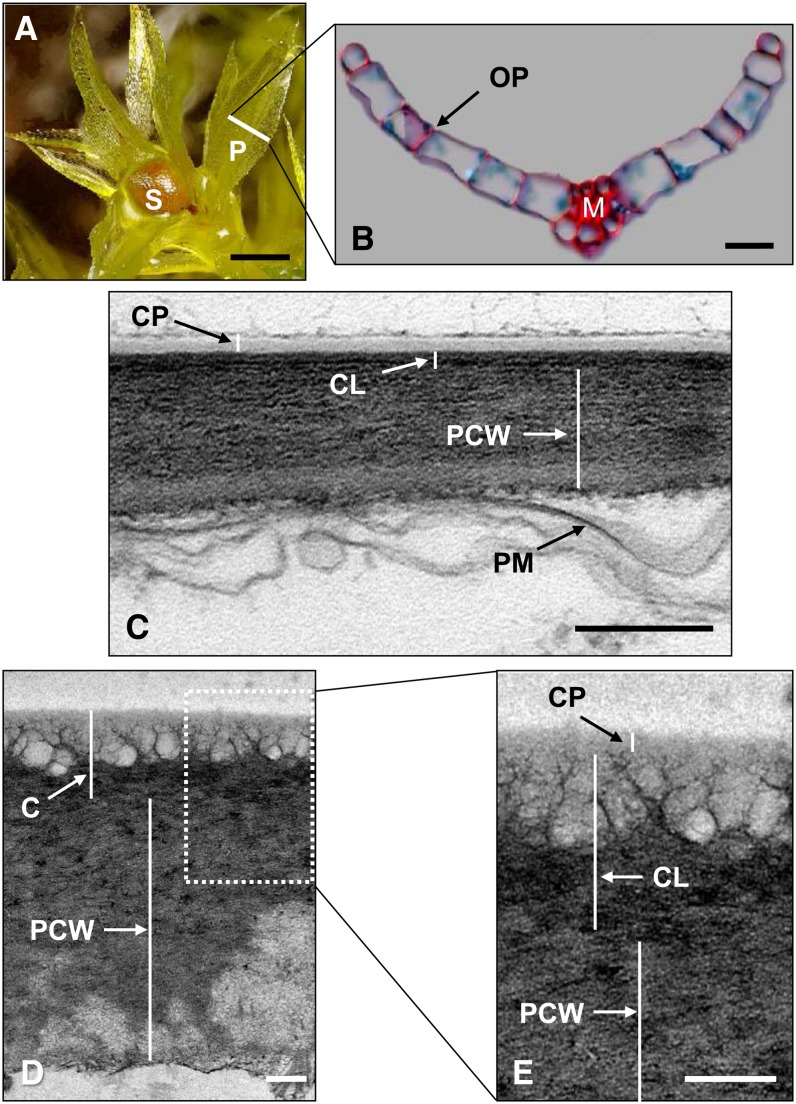
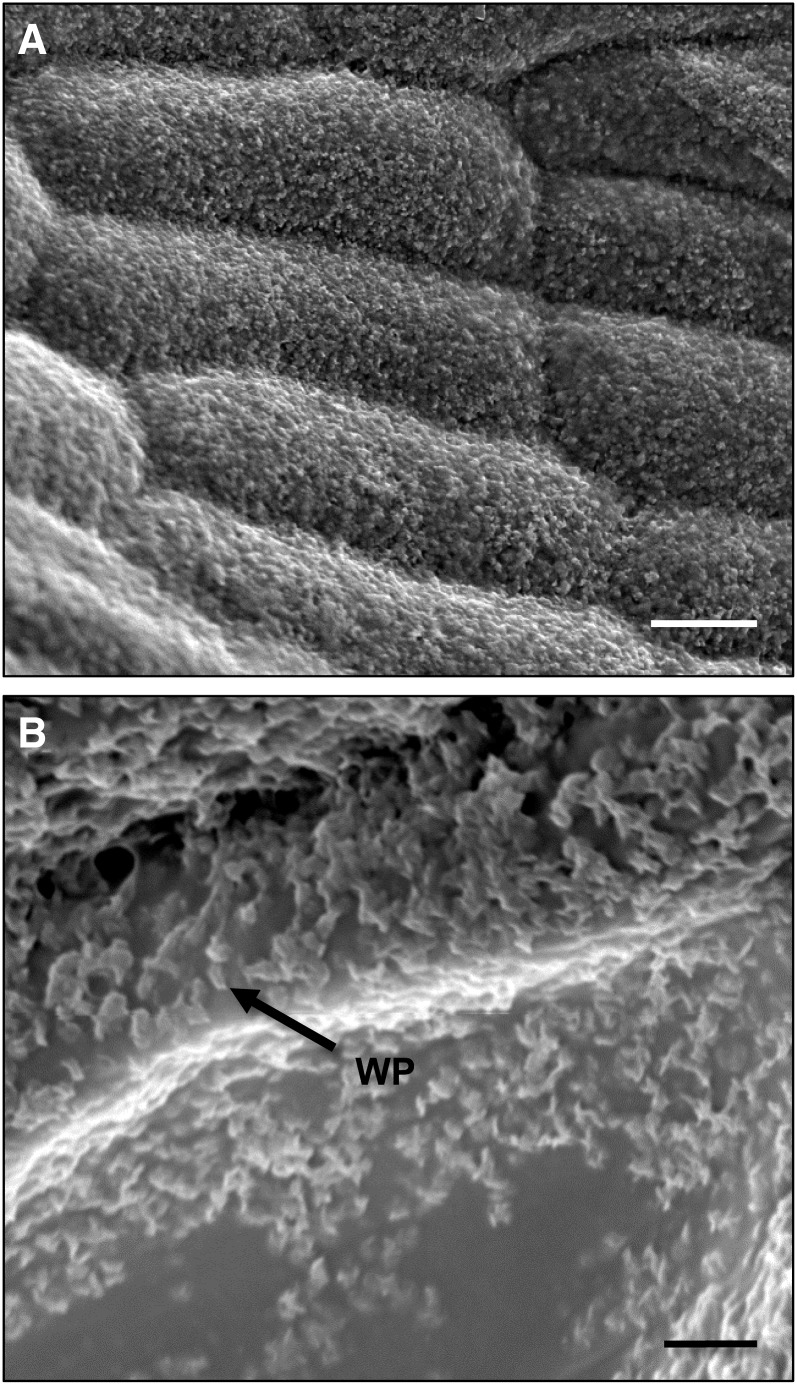
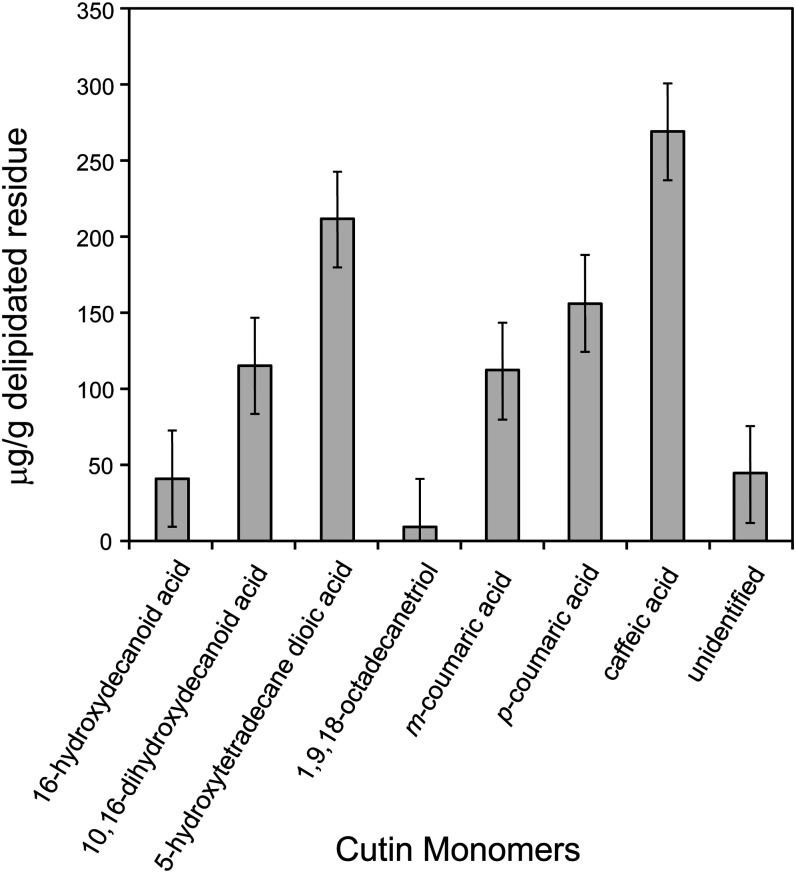
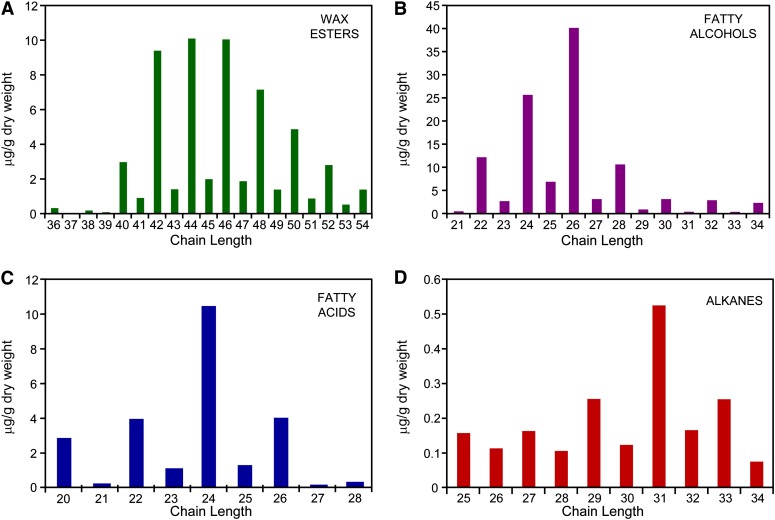
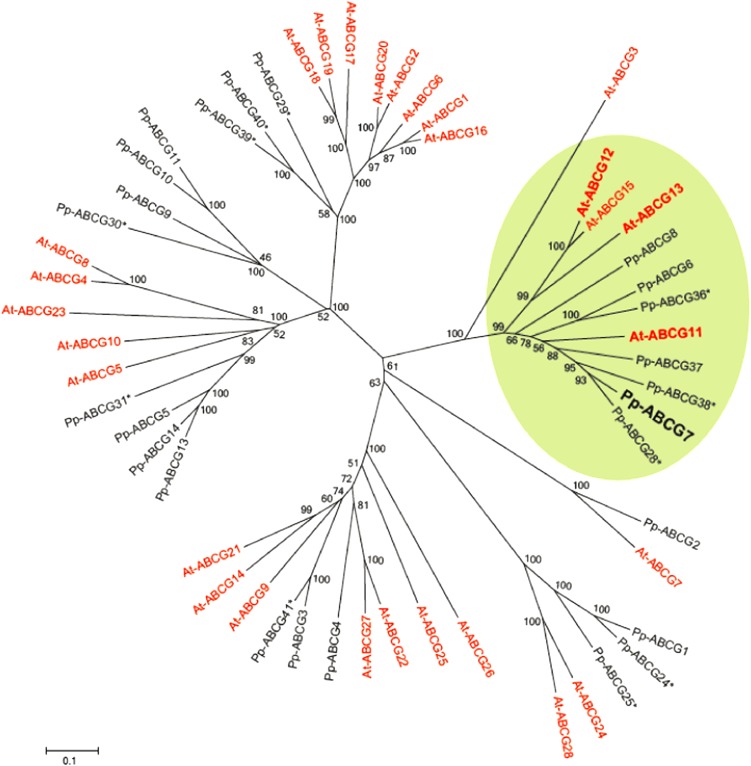
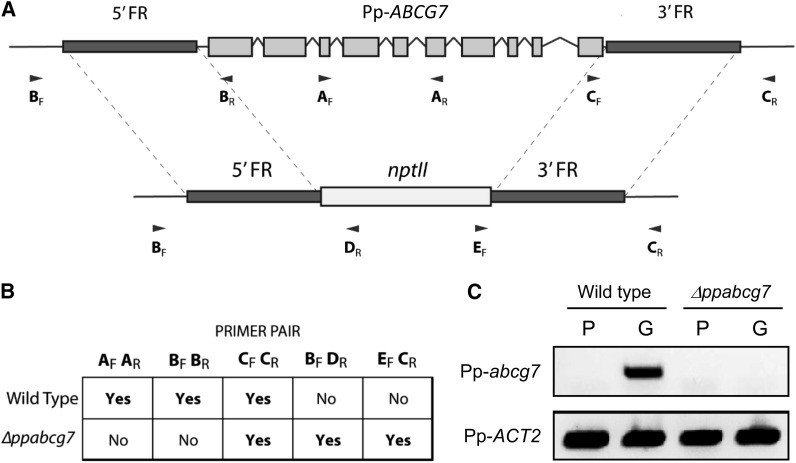
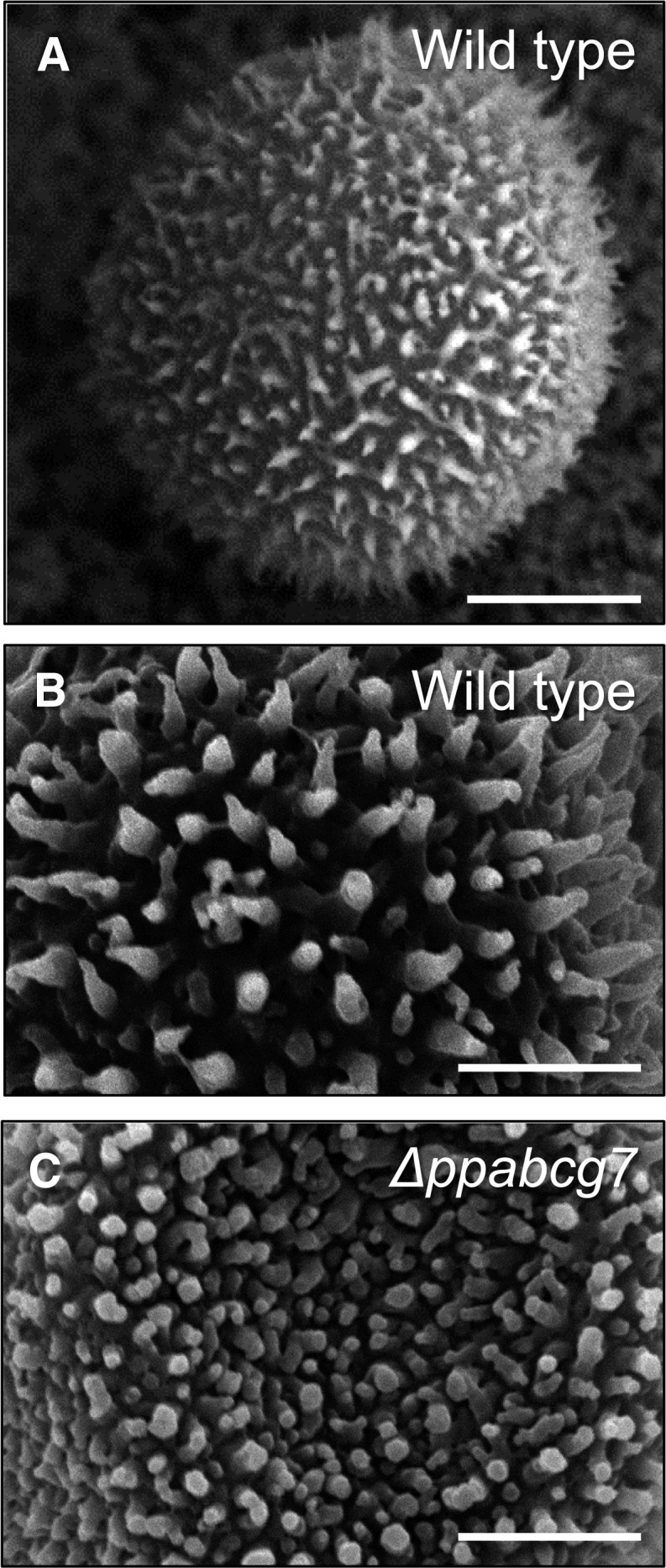
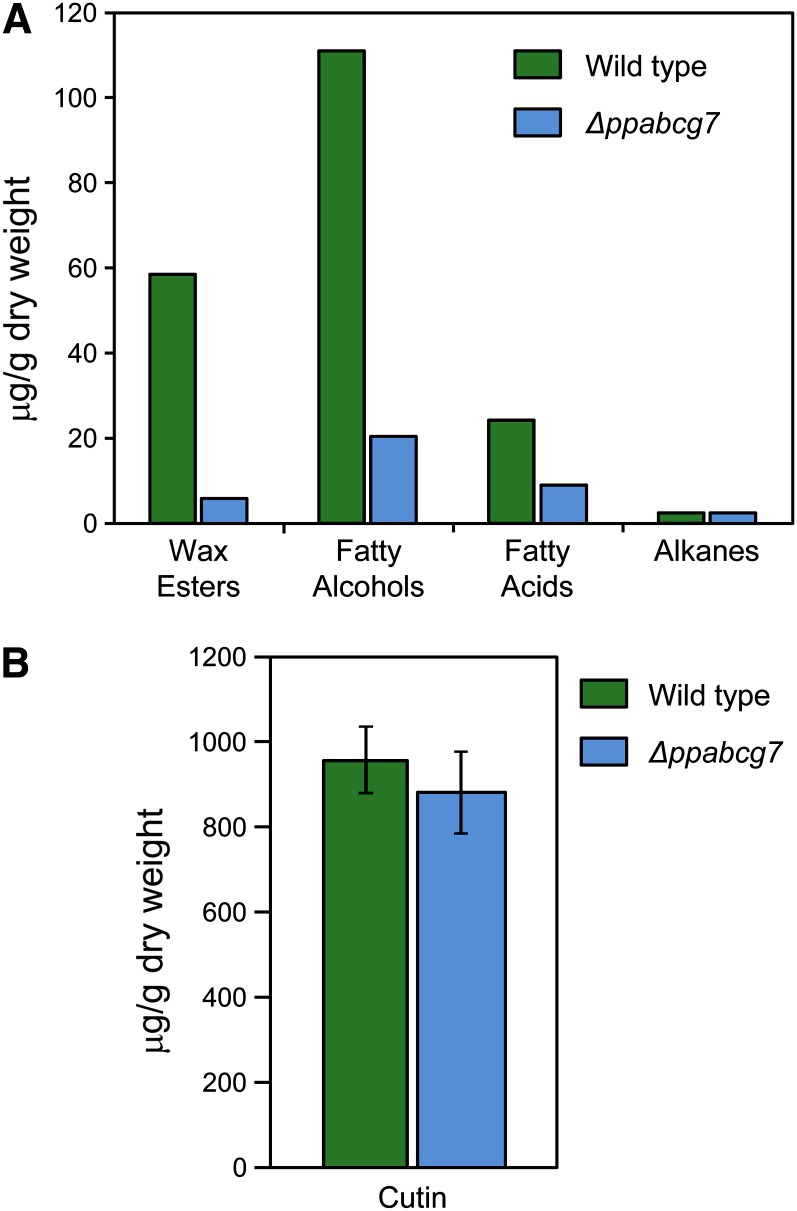
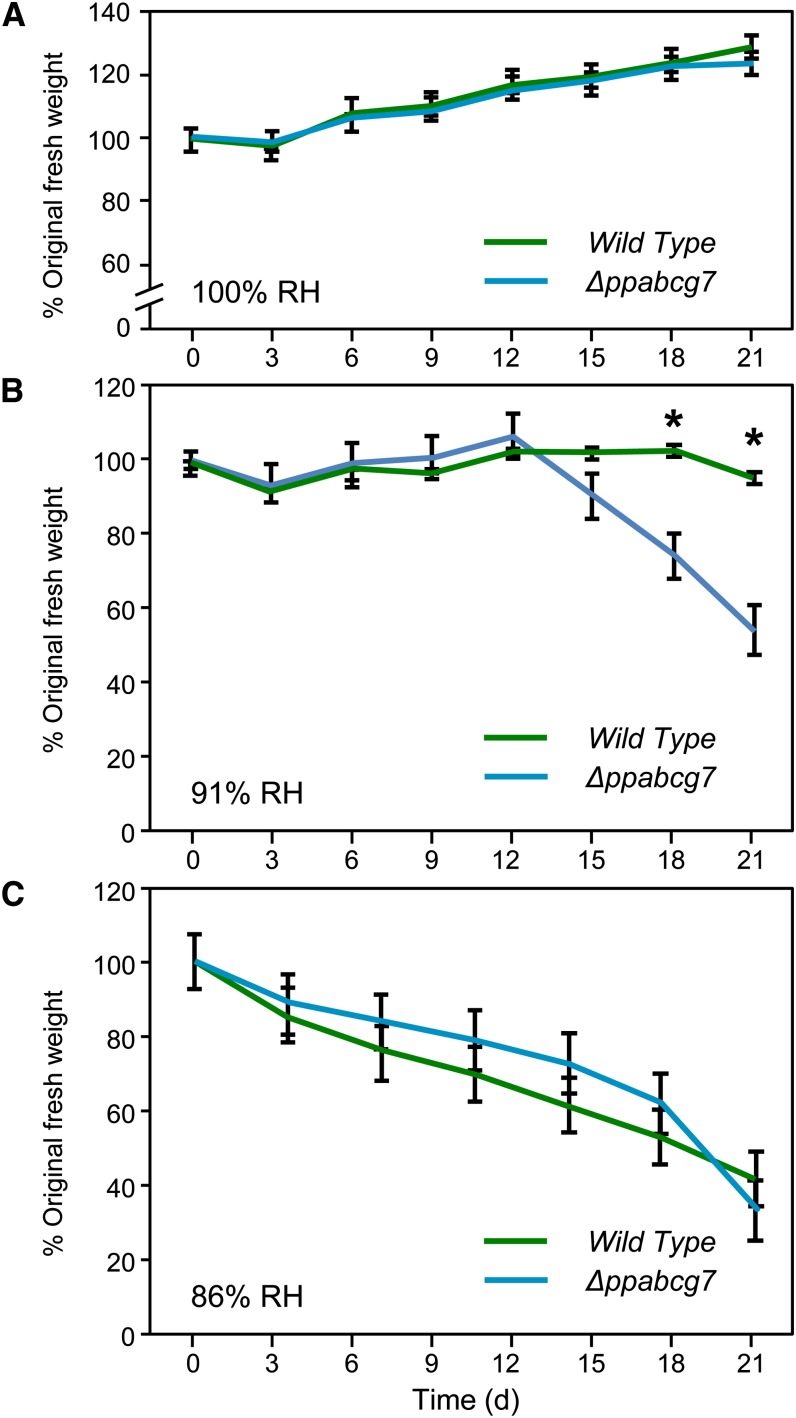
The plant cuticle is thought to be a critical evolutionary adaptation that allowed the first plants to colonize land, because of its key roles in regulating plant water status and providing protection from biotic and abiotic stresses. Much has been learned about cuticle composition and structure through genetic and biochemical studies of angiosperms, as well as underlying genetic pathways, but little is known about the cuticles of early diverging plant lineages. Here, we demonstrate that the moss Physcomitrella patens, an extant relative of the earliest terrestrial plants, has a cuticle that is analogous in both structure and chemical composition to those of angiosperms. To test whether the underlying cuticle biosynthetic pathways were also shared among distant plant lineages, we generated a genetic knockout of the moss ATP binding cassette subfamily G (ABCG) transporter Pp-ABCG7, a putative ortholog of Arabidopsis thaliana ABCG transporters involved in cuticle precursor trafficking. We show that this mutant is severely deficient in cuticular wax accumulation and has a reduced tolerance of desiccation stress compared with the wild type. This work provides evidence that the cuticle was an adaptive feature present in the first terrestrial plants and that the genes involved in their formation have been functionally conserved for over 450 million years.
Figures

References
-
- Baker E.A., Gaskin R.E. (1987). Composition of leaf epicuticular waxes of Pteridium subspecies. Phytochemistry 26: 2847–2848
-
- Barthlott W., Neinhuis C. (1997). Purity of the sacred lotus, or escape from contamination in biological surfaces. Planta 202: 1–8
-
- Beerling D.J., Franks P.J. (2009). Evolution of stomatal function in ‘lower’ land plants. New Phytol. 183: 921–925 - PubMed
-
- Beisson F., Li-Beisson Y., Pollard M. (2012). Solving the puzzles of cutin and suberin polymer biosynthesis. Curr. Opin. Plant Biol. 15: 329–337 - PubMed
-
- Bird D., Beisson F., Brigham A., Shin J., Greer S., Jetter R., Kunst L., Wu X., Yephremov A., Samuels L. (2007). Characterization of Arabidopsis ABCG11/WBC11, an ATP binding cassette (ABC) transporter that is required for cuticular lipid secretion. Plant J. 52: 485–498 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
