

Inefficient translation renders the Enterococcus faecalis fabK enoyl-acyl carrier protein reductase phenotypically cryptic
- PMID: 24163335
- PMCID: PMC3911128
- DOI: 10.1128/JB.01148-13
Inefficient translation renders the Enterococcus faecalis fabK enoyl-acyl carrier protein reductase phenotypically cryptic
Abstract
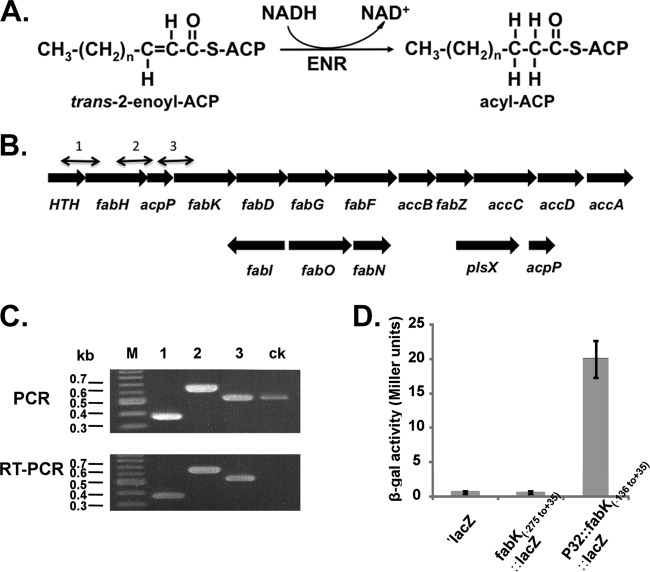
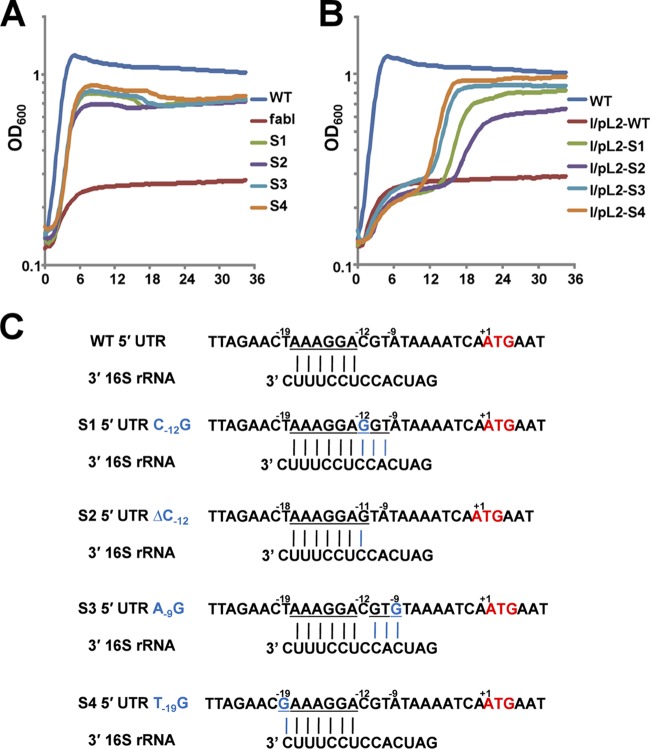
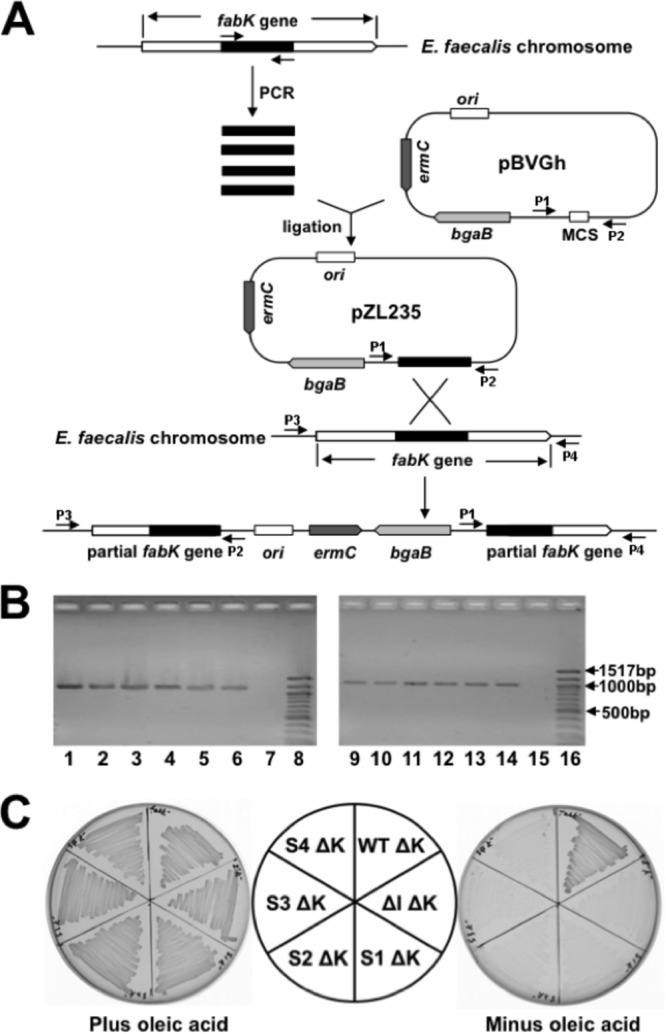
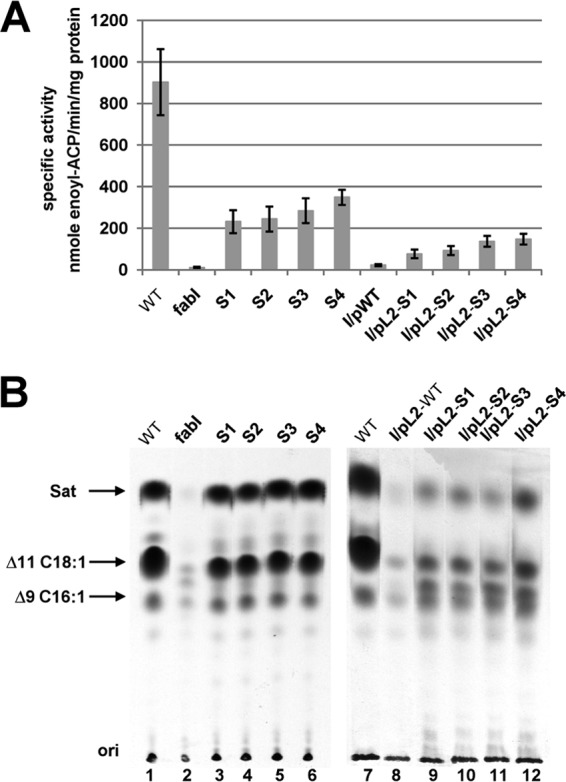
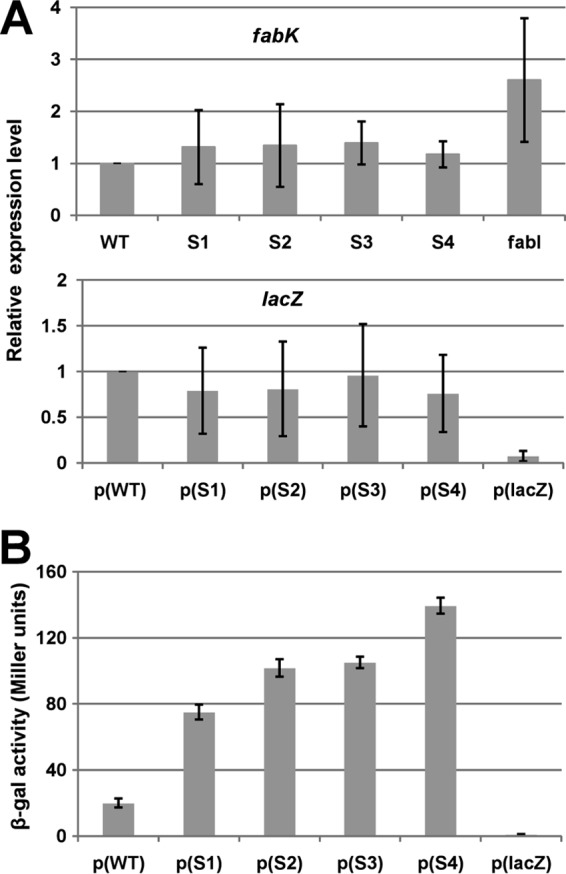
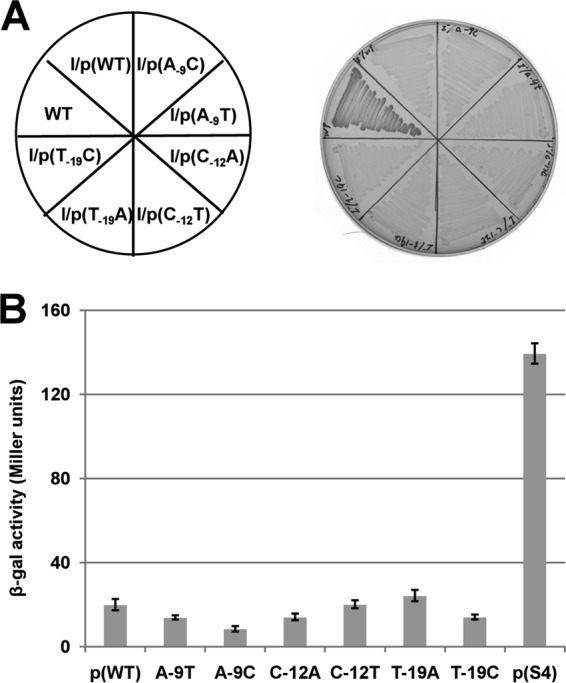
Enoyl-acyl carrier protein (ACP) reductase catalyzes the last step of the bacterial fatty acid elongation cycle. Enterococcus faecalis is unusual in that it encodes two unrelated enoyl-ACP reductases, FabI and FabK. We recently reported that deletion of the gene encoding FabI results in an unsaturated fatty acid (UFA) auxotroph despite the presence of fabK, a gene encoding a second fully functional enoyl-ACP reductase. By process of elimination, our prior report argued that poor expression was the reason that fabK failed to functionally replace FabI. We now report that FabK is indeed poorly expressed and that the expression defect is at the level of translation rather than transcription. We isolated four spontaneous mutants that allowed growth of the E. faecalis ΔfabI strain on fatty acid-free medium. Each mutational lesion (single base substitution or deletion) extended the fabK ribosome binding site. Inactivation of fabK blocked growth, indicating that the mutations acted only on fabK rather than a downstream gene. The mutations activated fabK translation to levels that supported fatty acid synthesis and hence cell growth. Furthermore, site-directed and random mutagenesis experiments showed that point mutations that resulted in increased complementarity to the 3' end of the 16S rRNA increased FabK translation to levels sufficient to support growth, whereas mutations that decreased complementarity blocked fabK translation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
