

Growth phase and pH influence peptide signaling for competence development in Streptococcus mutans
- PMID: 24163340
- PMCID: PMC3911236
- DOI: 10.1128/JB.00995-13
Growth phase and pH influence peptide signaling for competence development in Streptococcus mutans
Abstract
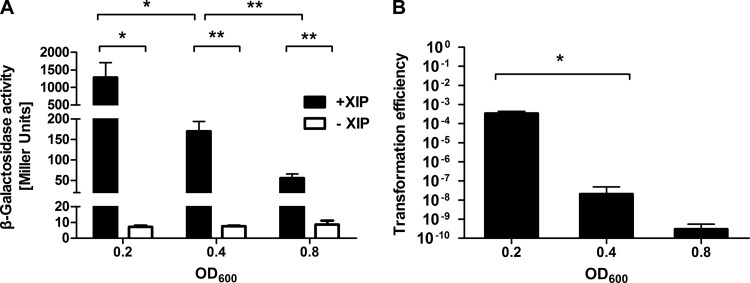
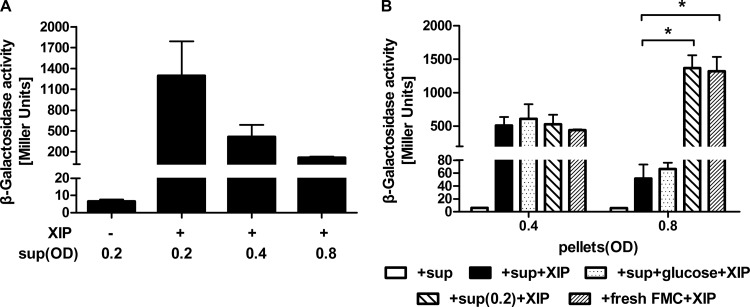
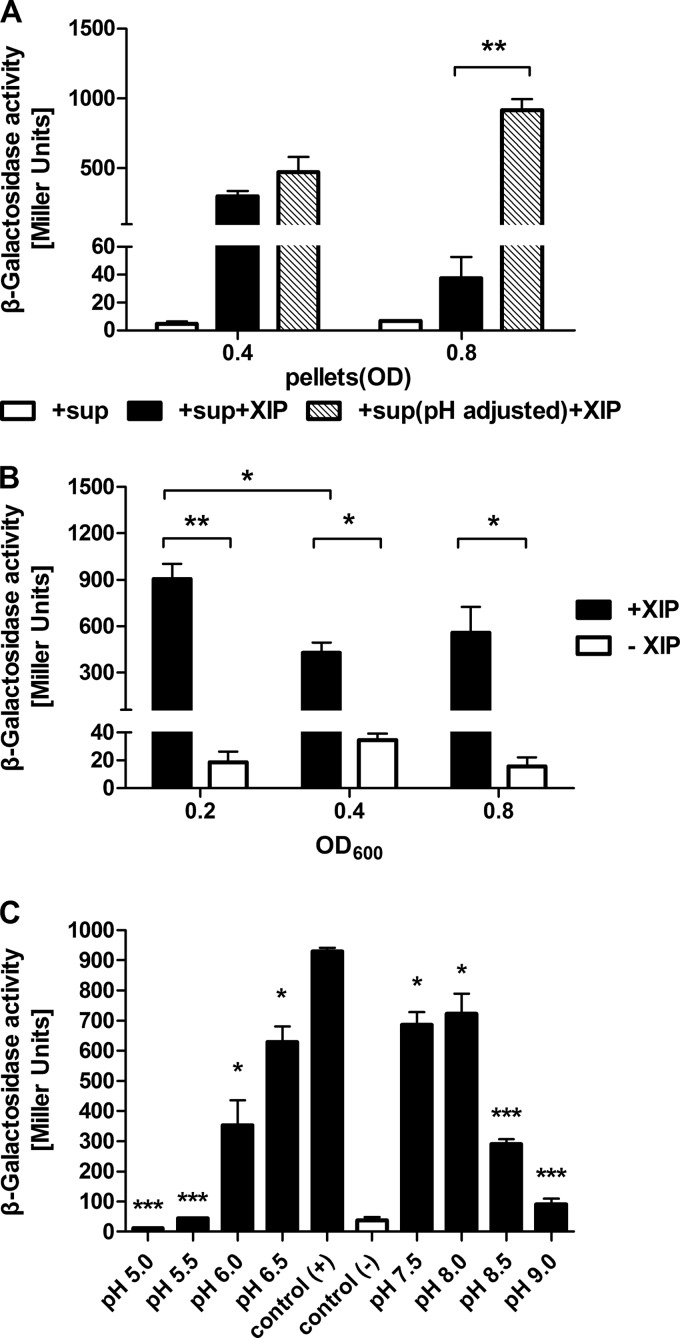
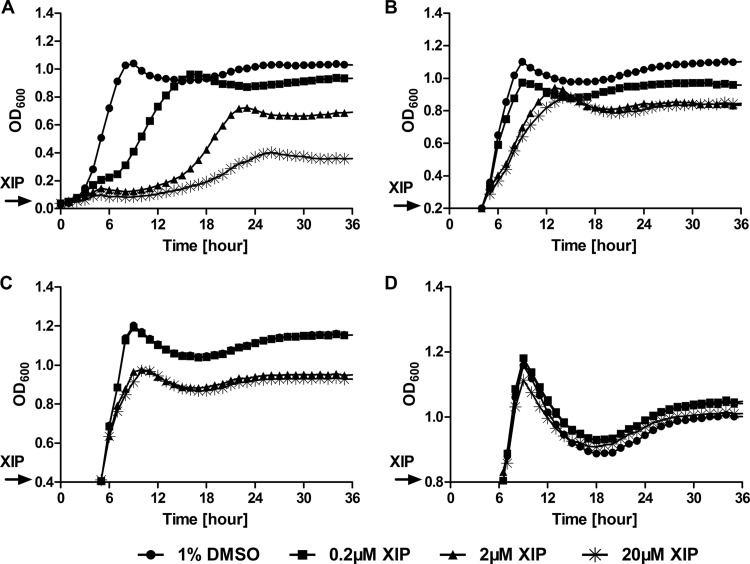
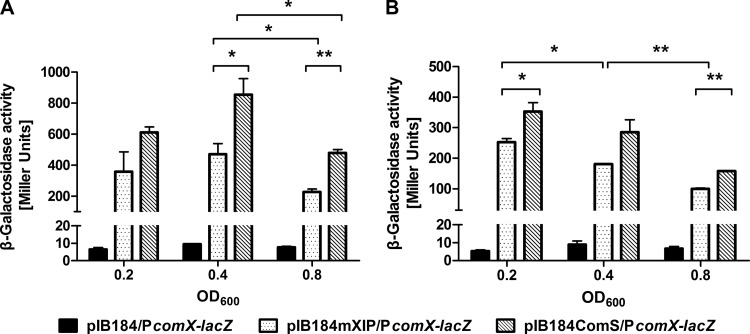
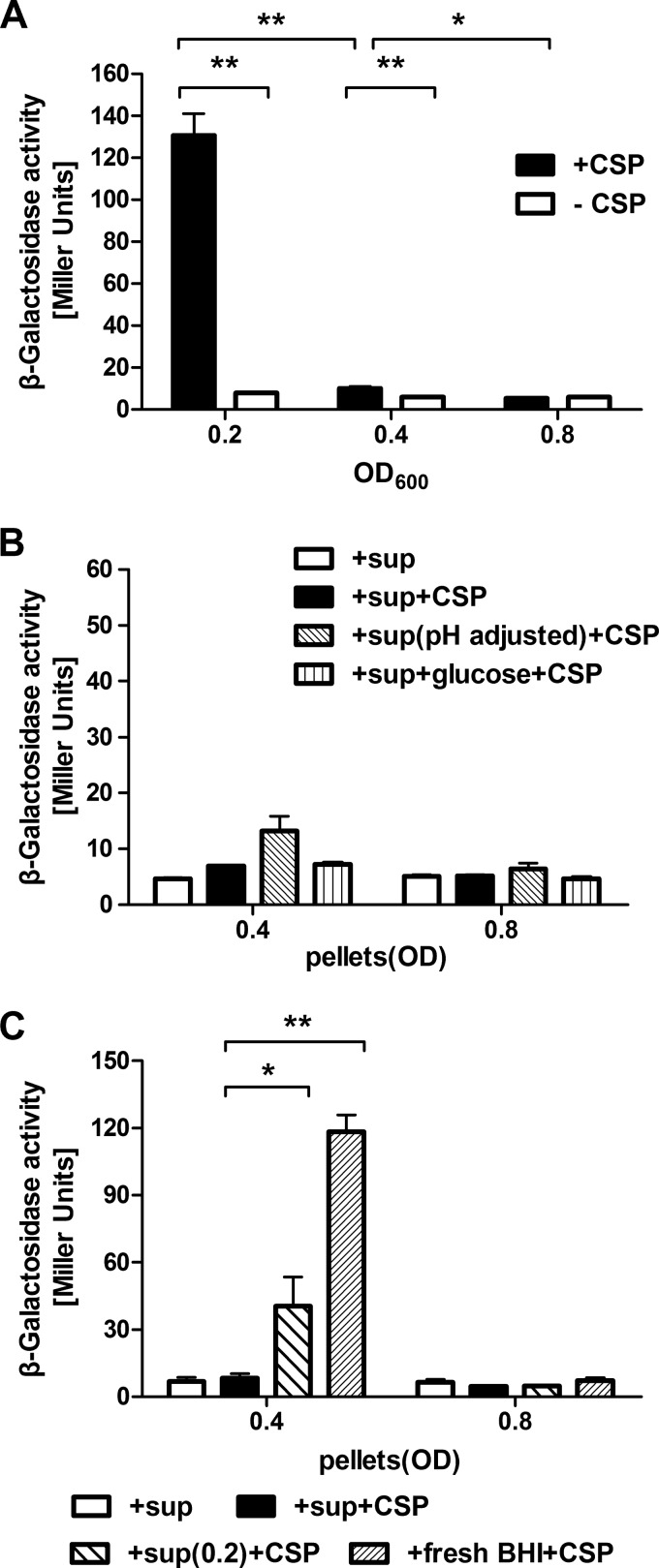
The development of competence by the dental caries pathogen Streptococcus mutans is mediated primarily through the alternative sigma factor ComX (SigX), which is under the control of multiple regulatory systems and activates the expression of genes involved in DNA uptake and recombination. Here we report that the induction of competence and competence gene expression by XIP (sigX-inducing peptide) and CSP (competence-stimulating peptide) is dependent on the growth phase and that environmental pH has a potent effect on the responses to XIP. A dramatic decline in comX and comS expression was observed in mid- and late-exponential-phase cells. XIP-mediated competence development and responses to XIP were optimal around a neutral pH, although mid-exponential-phase cells remained refractory to XIP treatment, and acidified late-exponential-phase cultures were resistant to killing by high concentrations of XIP. Changes in the expression of the genes for the oligopeptide permease (opp), which appears to be responsible for the internalization of XIP, could not entirely account for the behaviors observed. Interestingly, comS and comX expression was highly induced in response to endogenously overproduced XIP or ComS in mid-exponential-phase cells. In contrast to the effects of pH on XIP, competence induction and responses to CSP in complex medium were not affected by pH, although a decreased response to CSP in cells that had exited early-exponential phase was observed. Collectively, these results indicate that competence development may be highly sensitive to microenvironments within oral biofilms and that XIP and CSP signaling in biofilms could be spatially and temporally heterogeneous.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
