

Identification of the immunoproteasome as a novel regulator of skeletal muscle differentiation
- PMID: 24164898
- PMCID: PMC3911280
- DOI: 10.1128/MCB.00622-13
Identification of the immunoproteasome as a novel regulator of skeletal muscle differentiation
Abstract
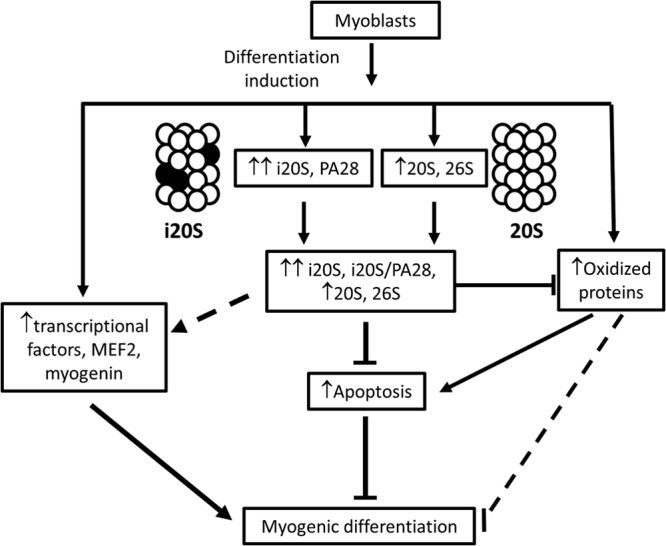
While many of the molecular details of myogenesis have been investigated extensively, the function of immunoproteasomes (i-proteasomes) in myogenic differentiation remains unknown. We show here that the mRNA of i-proteasome subunits, the protein levels of constitutive and inducible proteasome subunits, and the proteolytic activities of the 20S and 26S proteasomes were significantly upregulated during differentiation of skeletal muscle C2C12 cells. Knockdown of the i-proteasome catalytic subunit PSMB9 by short hairpin RNA (shRNA) decreased the expression of both PSMB9 and PSMB8 without affecting other catalytic subunits of the proteasome. PSMB9 knockdown and the use of i-proteasome-specific inhibitors both decreased 26S proteasome activities and prevented C2C12 differentiation. Inhibition of the i-proteasome also impaired human skeletal myoblast differentiation. Suppression of the i-proteasome increased protein oxidation, and these oxidized proteins were found to be more susceptible to degradation by exogenous i-proteasomes. Downregulation of the i-proteasome also increased proapoptotic proteins, including Bax, as well as cleaved caspase 3, cleaved caspase 9, and cleaved poly(ADP-ribose) polymerase (PARP), suggesting that impaired differentiation is likely to occur because of significantly increased apoptosis. These results demonstrate for the first time that i-proteasomes, independent of constitutive proteasomes, are critical for skeletal muscle differentiation of mouse C2C12 cells.
Figures







References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials