

Genome dynamics during experimental evolution
- PMID: 24166031
- PMCID: PMC4239992
- DOI: 10.1038/nrg3564
Genome dynamics during experimental evolution
Abstract
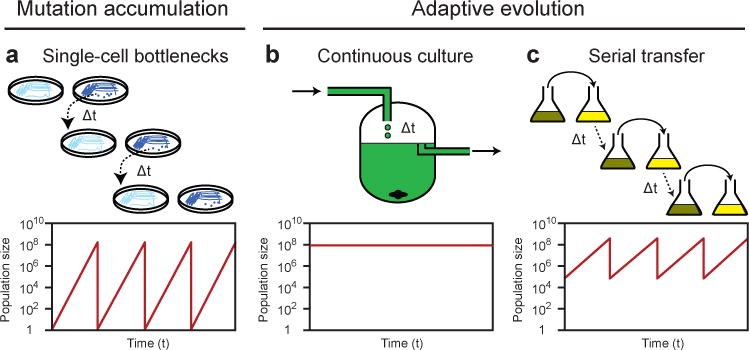
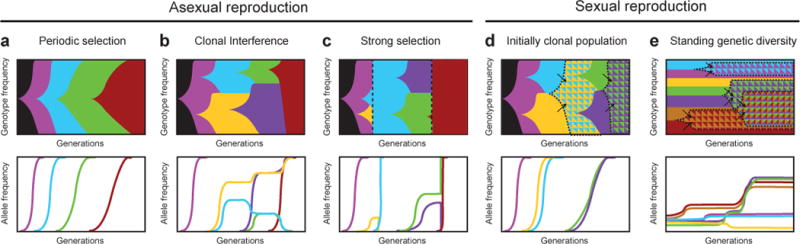
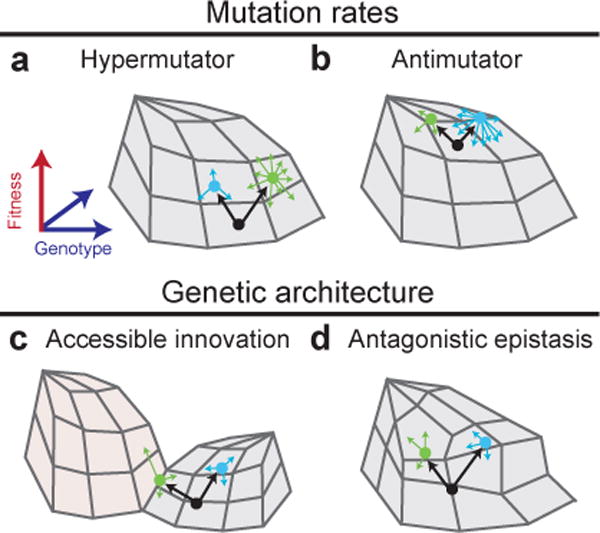
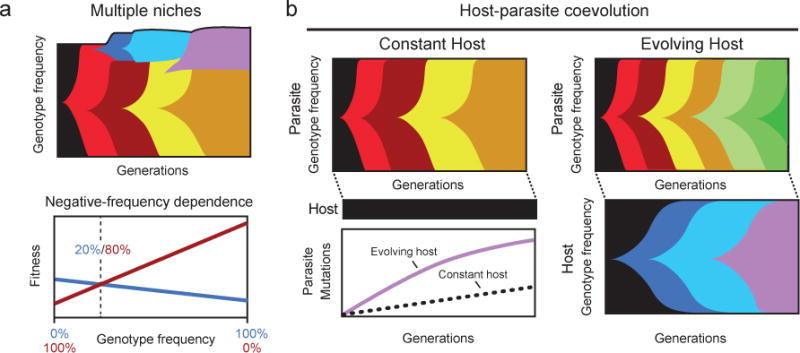
Evolutionary changes in organismal traits may occur either gradually or suddenly. However, until recently, there has been little direct information about how phenotypic changes are related to the rate and the nature of the underlying genotypic changes. Technological advances that facilitate whole-genome and whole-population sequencing, coupled with experiments that 'watch' evolution in action, have brought new precision to and insights into studies of mutation rates and genome evolution. In this Review, we discuss the evolutionary forces and ecological processes that govern genome dynamics in various laboratory systems in the context of relevant population genetic theory, and we relate these findings to evolution in natural populations.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
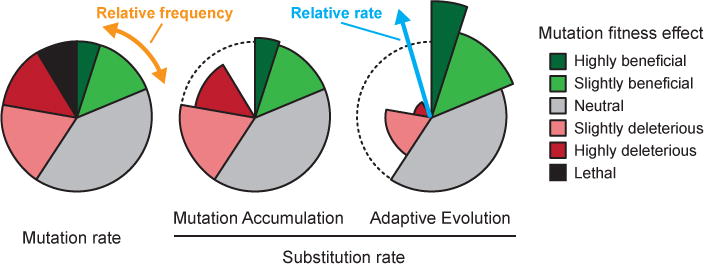
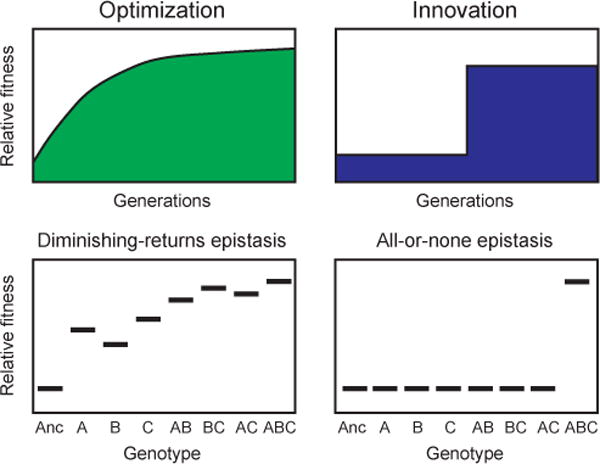
References
-
- Garland T, Rose MR, editors. Experimental Evolution: Concepts, Methods, and Applications of Selection Experiments. Univ. of California Press; 2009.
-
- Kawecki TJ, et al. Experimental evolution. Trends Ecol Evol. 2012;27:547–560. - PubMed
-
- Hartl DL, Clark AG. Principles of Population Genetics. Sinauer; Sunderland, MA: 2007.
-
- Mardis ER. Next-generation DNA sequencing methods. Annu Rev Genom Human Genet. 2008;9:387–402. - PubMed
-
- Schadt EE, Turner S, Kasarskis A. A window into third-generation sequencing. Hum Mol Genet. 2010;19:R227–R240. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
