

Social competition and selection in males and females
- PMID: 24167304
- PMCID: PMC3826203
- DOI: 10.1098/rstb.2013.0074
Social competition and selection in males and females
Abstract
During the latter half of the last century, evidence of reproductive competition between males and male selection by females led to the development of a stereotypical view of sex differences that characterized males as competitive and aggressive, and females as passive and choosy, which is currently being revised. Here, we compare social competition and its consequences for selection in males and females and argue that similar selection processes operate in both sexes and that contrasts between the sexes are quantitative rather than qualitative. We suggest that classifications of selection based on distinction between the form of competition or the components of fitness that are involved introduce unnecessary complexities and that the most useful approach in understanding the evolution and distribution of differences and similarities between the sexes is to compare the operation of selection in males and females in different reproductive systems.
Keywords: dominance status; mating systems; sex roles; sexual selection; social competition; social selection.
Figures


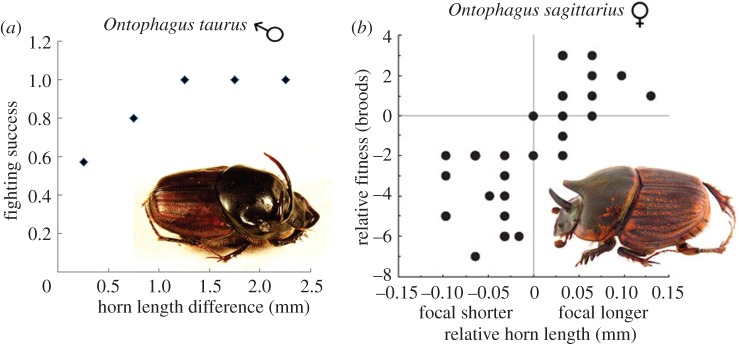
References
-
- Wilson EO. 1971. The insect societies. Cambridge, MA: Belknap Press
-
- Wilson EO. 1975. Sociobiology. Cambridge, MA: Harvard University Press
-
- Crook JH. 1972. Sexual selection, dimorphism and social organization in the primates. In Sexual selection and the descent of man 1871–1971 (ed. Campbell B.), pp. 231–281 Chicago, IL: Aldine
-
- West-Eberhard MJ. 1979. Sexual selection, social competition and evolution. Proc. R. Soc. Lond. B 123, 222–234
-
- Wynne-Edwards VC. 1962. Animal dispersion in relation to social behaviour. Edinburgh, UK: Oliver and Boyd
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources