

doi: 10.1021/nl403469r.
Epub 2013 Nov 13.
Nanopore-based identification of individual nucleotides for direct RNA sequencing
Affiliations
- PMID: 24171554
- PMCID: PMC3899427
- DOI: 10.1021/nl403469r
Item in Clipboard
Nanopore-based identification of individual nucleotides for direct RNA sequencing
Nano Lett.
2013.
Abstract
We describe a label-free ribobase identification method, which uses ionic current measurement to resolve ribonucleoside monophosphates or diphosphates in α-hemolysin protein nanopores containing amino-cyclodextrin adapters. The accuracy of base identification is further investigated through the use of a guanidino-modified adapter. On the basis of these findings, an exosequencing approach is envisioned in which a processive exoribonuclease (polynucleotide phosphorylase) presents sequentially cleaved ribonucleoside diphosphates to a nanopore.
Figures
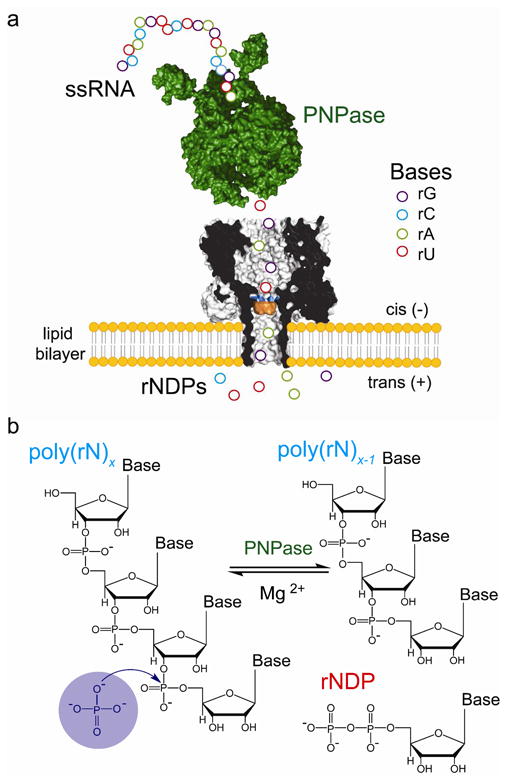
Detection of nucleotides cleaved from ssRNA by polynucleotide phosphorylase. (a) Schematic representation of a ssRNA oligonucleotide (circles) digested by polynucleotide phosphorylase (PNPase, green), one base at a time. The liberated nucleotides (rNDPs) are detected by the heptameric αHL pore (7AHL) equipped with a cyclodextrin adapter (am7βCD, orange). The mutant M113R (mutation highlighted in blue) was used in most of the experiments reported in this paper. In a functioning nanopore sequencer, the PNPase would be covalently attached to the αHL pore. (b) PNPase processively cleaves ssRNA substrates in the 3′-to-5′ direction by using inorganic phosphate (Pi) to attack the phosphodiester linkage nearest the 3′ terminus to release rNDPs.
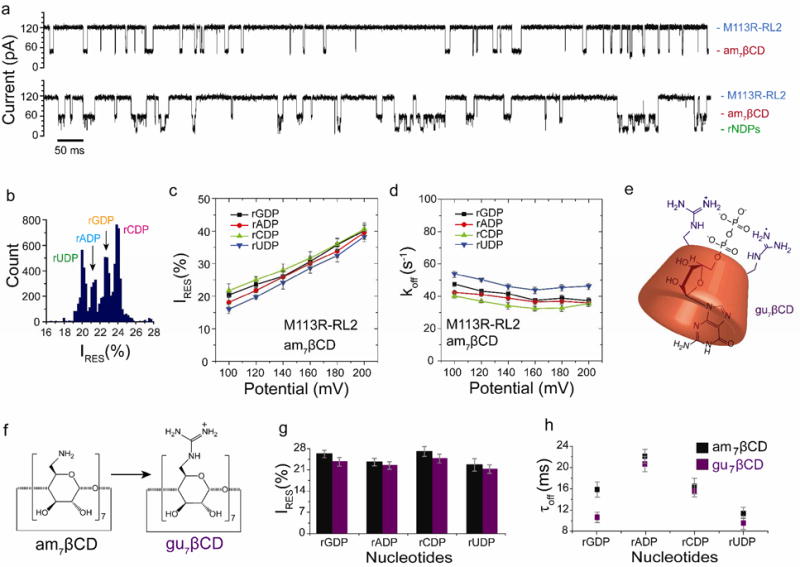
Nucleoside diphosphate discrimination with non-covalently attached cyclodextrin adapters. (a) Single-channel recordings from the homoheptameric αHL M113R-RL2 pore showing am7βCD binding (top) and rNDP detection with the am7βCD adapter (bottom). (b) Corresponding residual current (IRES%) histogram. Data were acquired in 1.2 M KCl, 25 mM Tris-HCl, pH 6.0, at +120 mV in the presence of 80 μM am7βCD (trans), 10 μM rGDP, 10 μM rADP, 10 μM rCDP and 10 μM rUDP (all cis). The results displayed are from a typical experiment. (c) Variation of residual currents (IRES%) with applied potential for each rNDP detected with the M1 13R-RL2•am7βCD pore. (d) Variation of koff with applied potential for each rNDP detected with the M1 13R-RL2•7βCD pore. Values of koff were determined by using koff = 1/τoff, where τoff is the mean dwell time for each rNDP in the pore. (e) Schematic representation of the complex of rGDP with gu7βCD, created using ChemBioDraw software (version 12.02). (f) Conversion of am7βCD to gu7βCD. For clarity, only two of the seven guanidino groups are shown. (g) Comparison of residual currents (IRES%) for each rNDP detected with the am7βCD and gu7βCD cyclodextrins within the αHL M113R-RL2 pore (n = 22). (h) mean dwell times (τoff) for each rNDP detected with the am7βCD and gu7βCD cyclodextrins within the αHL M113R-RL2 pore (n = 22).
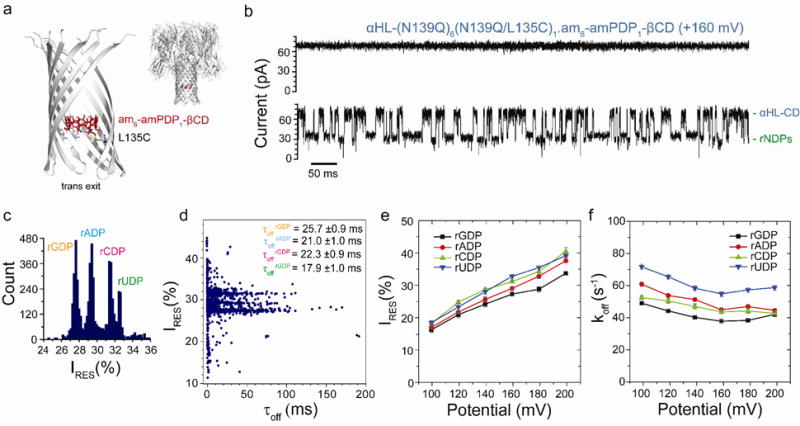
rNDP interaction with the αHL-(N139Q)6(N139Q/L135C)1.am6-amPDP1-βCD pore. (a) Structure of the αHL-(N139Q);6(N139Q/L135C)1.am6-amPDP1- βCD pore (cartoon view). The enlarged view shows a close-up of the β barrel with two subunits omitted. The cyclodextrin (am6-amPDP1-βCD) was covalently attached through a disulfide bond to position 135 in one of the seven subunits, which had been mutated to Cys (red, sticks model)(b) Single-channel recording from the αHL-(N139Q)6(N139Q/L135C)1.am6-amPDP1-βCD pore showing continuous rNDP detection. (c) Corresponding residual current (IRES%) histogram. Data were acquired in 1 M KCl, 25 mM Tris-HCl, pH 7.5, at +160 mV in the presence of 10 μM rGDP, 10 μM rADP, 10 μM rCDP and 10 μM rUDP (all cis). The results displayed are from a typical experiment. (d) Scatter plot showing IRES% and dwell times (τoff) for rNDP binding events to the αHL-(N139Q)6(N139Q/L135C)1.am6-amPDP1-βCD pore as seen in the current trace in ‘b’. (e) Variation of residual currents (IRES%) with applied potential for each rNDP detected with the αHL-(N139Q)6(N139Q/L135C)1.am6-amPDP1βCD pore. (f) Variation of k Values of koff with applied potential for each rNDP detected with the αHL-(N139Q)6(N139Q/L135C)1.am6-amPDP1-βCD pore. Values of koff were determined by using koff = 1/τoff where τoff is the mean dwell time of the rNDP in the pore.
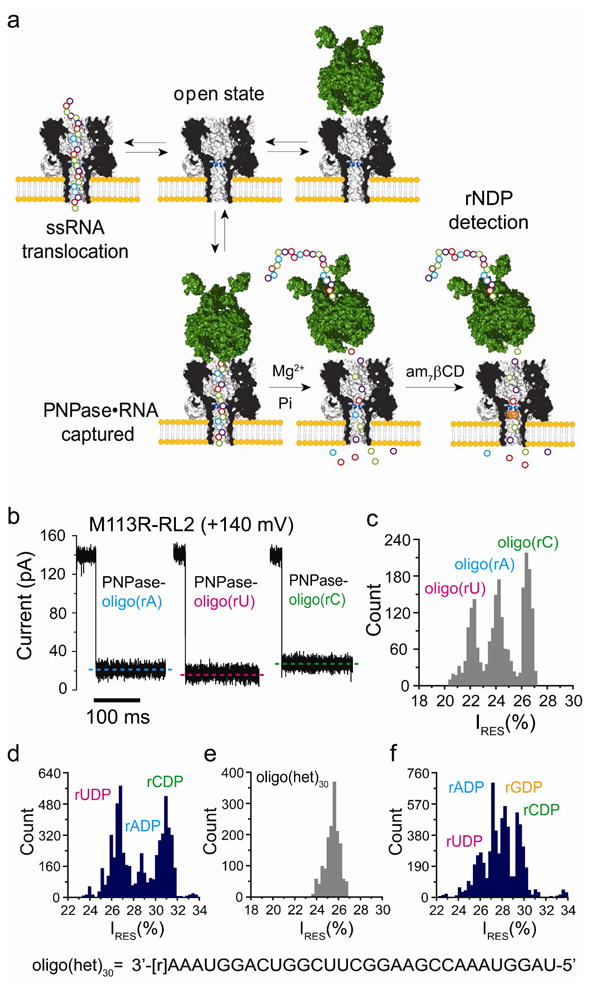
Detection of nucleosides diphosphates cleaved from ssRNA by PNPase. (a) Upon the addition of ssRNA, without the enzyme, current blockades are caused by brief ssRNA translocation events. In the presence of PNPase, additional blockades with longer residence times and/or amplitudes that differ from those of the translocation events are observed when PNPase•RNA complexes are captured. After the addition of Mg2+ and inorganic phosphate (Pi), followed by the addition of 7βCD (trans), cleaved nucleotides are detected through their interactions with the CD adapter. (b) Current traces for the capture of homopolymeric ssRNAs bound to PNPase by the αHL M113R-RL2 pore at +140 mV. (c) Histogram of the residual current levels (IRES%) after capture of homopolymers ssRNAs bound to PNPase by the αHL M113R-RL2 pore at +140 mV. (d) Histogram of the residual current levels (IRES%) for the rNDP binding events with am7βCD (trans). The rNDPs were cleaved by PNPase from the RNA homopolymers, oligo(rA)30, oligo(rC)30 and oligo(rU)30. e) Histogram of the residual current levels (IRES%) after capture of a PNPase-bound hetero-oligomeric ssRNA (oligo(het)30: 3′-[r]AAAUGGACUGGCUUCGGAAGCCAAAUGGAU-5′) inside the αHL M113R-RL2 pore at +120 mV. (f) Histogram of the residual current levels (IRES%) for rNDP binding events in the presence of am7β CD(trans). The rNDPs were cleaved by PNPase from the hetero-oligomeric ssRNA, oligo(het)30(Figure S11).
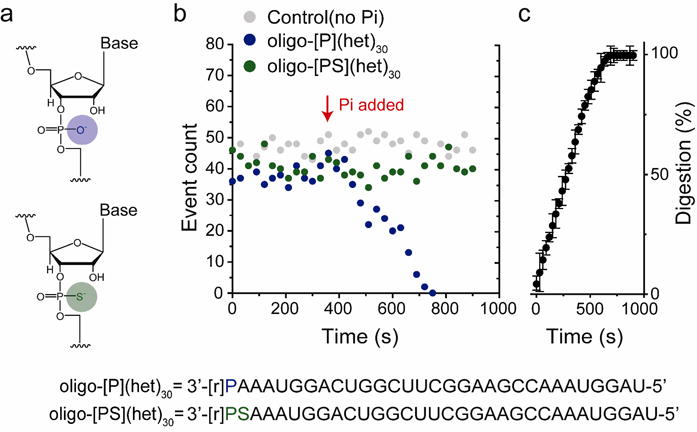
Monitoring PNPase digestion of ssRNA with a nanopore. (a) Chemical structures of the phosphodiester linkage in oligo-[P](het)30 (top, blue circle) and the phosphorothioate linkage in oligo-[PS](het)30 (bottom, green circle), which is not hydrolyzed by PNPase. (Below) Sequences of the hetero-oligomeric oligonucleotides. (b) The PNPase bound oligo-[P](het)30 ssRNA capture rate in the M113R-RL2 nanopore as a function of time. After the addition of 10 mM Pi (red arrow), the event rate sharply declines for the PNPase-oligo-[P](het)30. In contrast, the event rate is maintained for PNPase-oligo-[PS](het)30. (c) Time course for oligo-[P](het)30 ssRNA digestion by PNPase (n = 3). The plot was constructed by collecting capture events in a 30 s segment (from t= 0 s to 900 s), in the presence of Pi and Mg2+.
References
-
- Bayley H. Current Opinion in Chemical Biology. 2006;10(6):628–637. - PubMed
-
- Branton D, Deamer DW, Marziali A, Bayley H, Benner SA, Butler T, Di Ventra M, Garaj S, Hibbs A, Huang X, Jovanovich SB, Krstic PS, Lindsay S, Ling XS, Mastrangelo CH, Meller A, Oliver JS, Pershin YV, Ramsey JM, Riehn R, Soni GV, Tabard-Cossa V, Wanunu M, Wiggin M, Schloss JA. Nat Biotech. 2008;26(10):1146–1153. - PMC - PubMed
-
- Muthukumar M. Annual Review of Biophysics and Biomolecular Structure. 2007;36(1):435–450. - PubMed
-
- Song LZ, Hobaugh MR, Shustak C, Cheley S, Bayley H, Gouaux JE. Science. 1996;274(5294):1859–1866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
