

Characterization of a Brg1 hypomorphic allele demonstrates that genetic and biochemical activity are tightly correlated
- PMID: 24172864
- PMCID: PMC3962535
- DOI: 10.4161/epi.26879
Characterization of a Brg1 hypomorphic allele demonstrates that genetic and biochemical activity are tightly correlated
Abstract
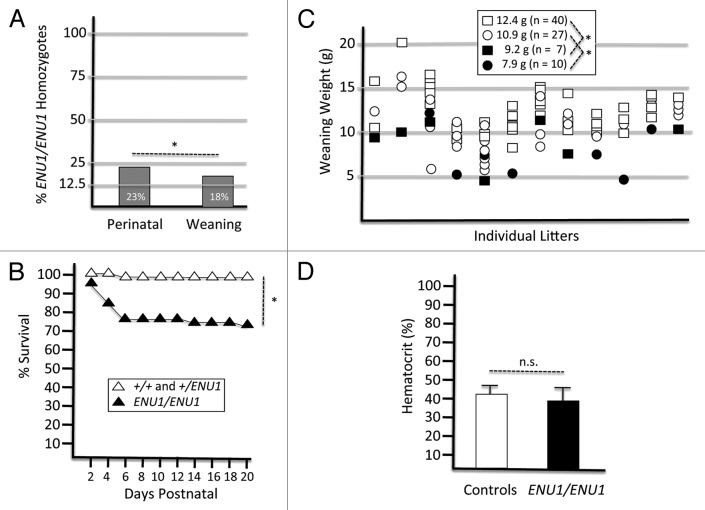
Mammalian SWI/SNF-related complexes are recruited to the promoters of numerous target genes, and the BRG1 catalytic subunit confers ATPase activity necessary to slide or evict nucleosomes and to regulate transcription. Based on gene-targeting experiments in mice, BRG1 is essential for early embryonic development. However, Brg1 null mutants have provided limited insight into gene-dosage considerations and structure-function relationships. To extend our knowledge of BRG1 function, we describe the genetic and biochemical characteristics of an ENU-induced hypomorphic mutation that encodes a protein with a single amino-acid substitution (E1083G) within the bilobal ATPase/chromatin-remodeling domain. Brg1(ENU1/ENU1) mice have ~50% genetic activity and survive embryogenesis but exhibit a postnatal developmental phenotype associated with runting and incompletely penetrant lethality. The E1083G mutant protein is stable, and experiments with recombinant FLAG-tagged BRG1 proteins demonstrated that it retains full ATPase activity. Yet the biochemical activity of the mutant protein is diminished to ~50% of normal in chromatin-remodeling assays. Consistent with these findings, the E1083G substitution is predicted to disrupt a structurally conserved α-helix within the lobe that participates in DNA translocation but does not contain the ATPase catalytic site. We propose that this α-helix participates in the DNA translocation cycle by mechanistically linking DNA interaction surfaces at the DNA entry/anchor point to those within the Helicase C domain of lobe 2 of the bilobal ATPase motor. Taken together, these results demonstrate that BRG1 genetic and biochemical activities are tightly correlated. They also indicate that BRG1 ATPase activity is necessary but not sufficient for chromatin remodeling.
Keywords: ATPase; BRG1 E1083G; chromatin remodeling; hypomorph; runting.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous