

Staphylococcus δ-toxin induces allergic skin disease by activating mast cells
- PMID: 24172897
- PMCID: PMC4090780
- DOI: 10.1038/nature12655
Staphylococcus δ-toxin induces allergic skin disease by activating mast cells
Abstract
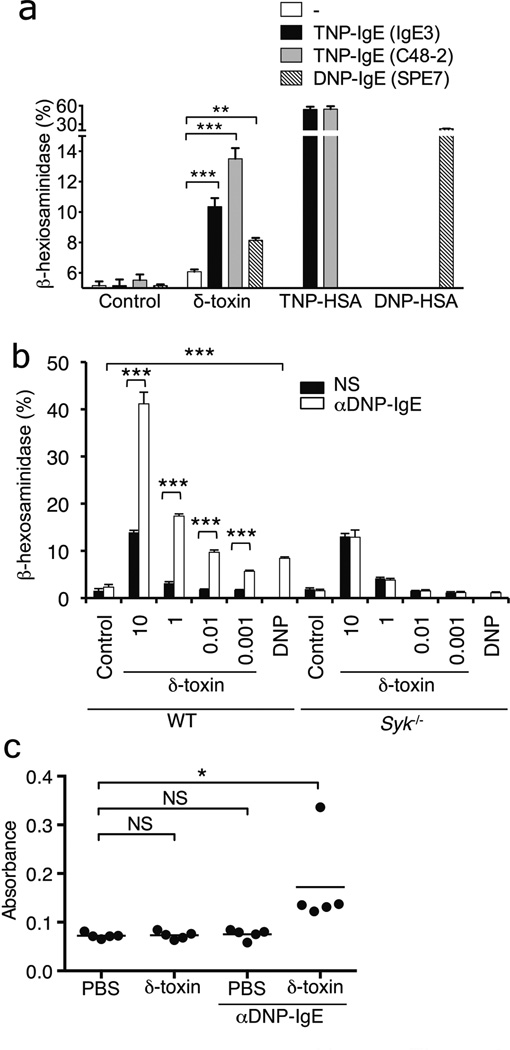
Atopic dermatitis is a chronic inflammatory skin disease that affects 15-30% of children and approximately 5% of adults in industrialized countries. Although the pathogenesis of atopic dermatitis is not fully understood, the disease is mediated by an abnormal immunoglobulin-E immune response in the setting of skin barrier dysfunction. Mast cells contribute to immunoglobulin-E-mediated allergic disorders including atopic dermatitis. Upon activation, mast cells release their membrane-bound cytosolic granules leading to the release of several molecules that are important in the pathogenesis of atopic dermatitis and host defence. More than 90% of patients with atopic dermatitis are colonized with Staphylococcus aureus in the lesional skin whereas most healthy individuals do not harbour the pathogen. Several staphylococcal exotoxins can act as superantigens and/or antigens in models of atopic dermatitis. However, the role of these staphylococcal exotoxins in disease pathogenesis remains unclear. Here we report that culture supernatants of S. aureus contain potent mast-cell degranulation activity. Biochemical analysis identified δ-toxin as the mast cell degranulation-inducing factor produced by S. aureus. Mast cell degranulation induced by δ-toxin depended on phosphoinositide 3-kinase and calcium (Ca(2+)) influx; however, unlike that mediated by immunoglobulin-E crosslinking, it did not require the spleen tyrosine kinase. In addition, immunoglobulin-E enhanced δ-toxin-induced mast cell degranulation in the absence of antigen. Furthermore, S. aureus isolates recovered from patients with atopic dermatitis produced large amounts of δ-toxin. Skin colonization with S. aureus, but not a mutant deficient in δ-toxin, promoted immunoglobulin-E and interleukin-4 production, as well as inflammatory skin disease. Furthermore, enhancement of immunoglobulin-E production and dermatitis by δ-toxin was abrogated in Kit(W-sh/W-sh) mast-cell-deficient mice and restored by mast cell reconstitution. These studies identify δ-toxin as a potent inducer of mast cell degranulation and suggest a mechanistic link between S. aureus colonization and allergic skin disease.
Figures



References
-
- Williams H, Flohr C. How epidemiology has challenged 3 prevailing concepts about atopic dermatitis. J Allergy Clin Immunol. 2006;118:209–213. - PubMed
-
- Liu FT, Goodarzi H, Chen HY. IgE, mast cells, and eosinophils in atopic dermatitis. Clin Rev Allergy Immunol. 2011;41:298–310. - PubMed
-
- Rudikoff D, Lebwohl M. Atopic dermatitis. Lancet. 1998;351:1715–1721. - PubMed
Supplementary References
-
- Bae T, Schneewind O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid. 2006;55:58–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
