

Apolipoprotein E codetermines tissue tropism of hepatitis C virus and is crucial for viral cell-to-cell transmission by contributing to a postenvelopment step of assembly
- PMID: 24173232
- PMCID: PMC3911621
- DOI: 10.1128/JVI.01815-13
Apolipoprotein E codetermines tissue tropism of hepatitis C virus and is crucial for viral cell-to-cell transmission by contributing to a postenvelopment step of assembly
Abstract
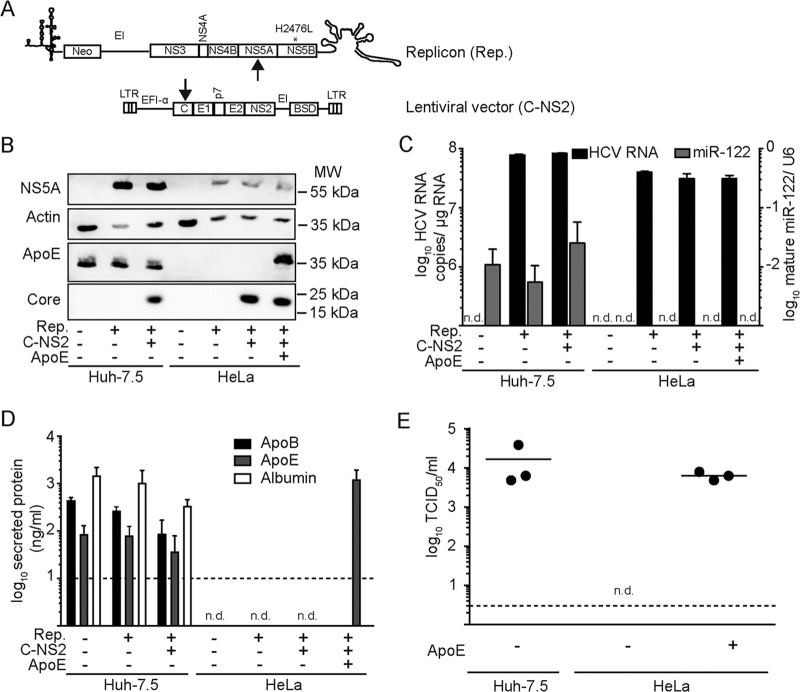
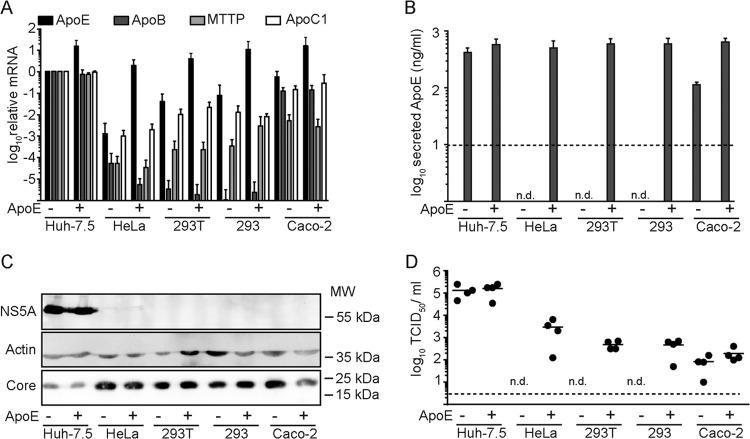
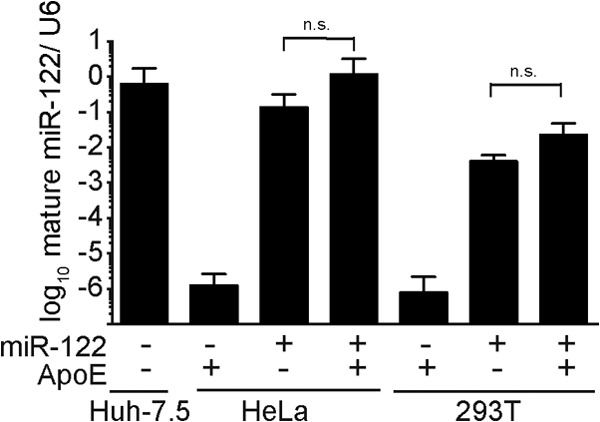
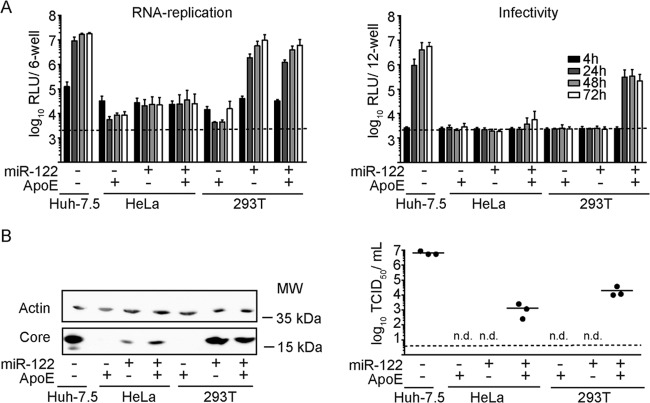
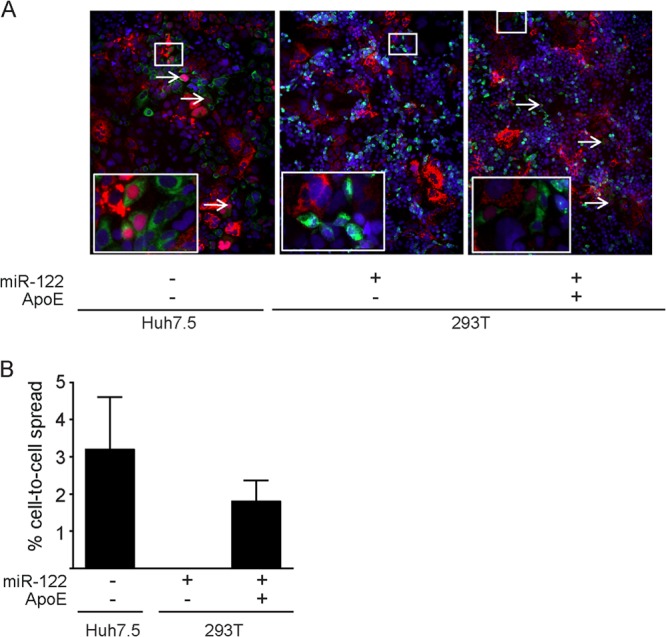
Hepatitis C virus (HCV) predominantly infects human hepatocytes, although extrahepatic virus reservoirs are being discussed. Infection of cells is initiated via cell-free and direct cell-to-cell transmission routes. Cell type-specific determinants of HCV entry and RNA replication have been reported. Moreover, several host factors required for synthesis and secretion of lipoproteins from liver cells, in part expressed in tissue-specific fashion, have been implicated in HCV assembly. However, the minimal cell type-specific requirements for HCV assembly have remained elusive. Here we report that production of HCV trans-complemented particles (HCVTCP) from nonliver cells depends on ectopic expression of apolipoprotein E (ApoE). For efficient virus production by full-length HCV genomes, microRNA 122 (miR-122)-mediated enhancement of RNA replication is additionally required. Typical properties of cell culture-grown HCV (HCVcc) particles from ApoE-expressing nonliver cells are comparable to those of virions derived from human hepatoma cells, although specific infectivity of virions is modestly reduced. Thus, apolipoprotein B (ApoB), microsomal triglyceride transfer protein (MTTP), and apolipoprotein C1 (ApoC1), previously implicated in HCV assembly, are dispensable for production of infectious HCV. In the absence of ApoE, release of core protein from infected cells is reduced, and production of extracellular as well as intracellular infectivity is ablated. Since envelopment of capsids was not impaired, we conclude that ApoE acts after capsid envelopment but prior to secretion of infectious HCV. Remarkably, the lack of ApoE also abrogated direct HCV cell-to-cell transmission. These findings highlight ApoE as a host factor codetermining HCV tissue tropism due to its involvement in a late assembly step and viral cell-to-cell transmission.
Figures
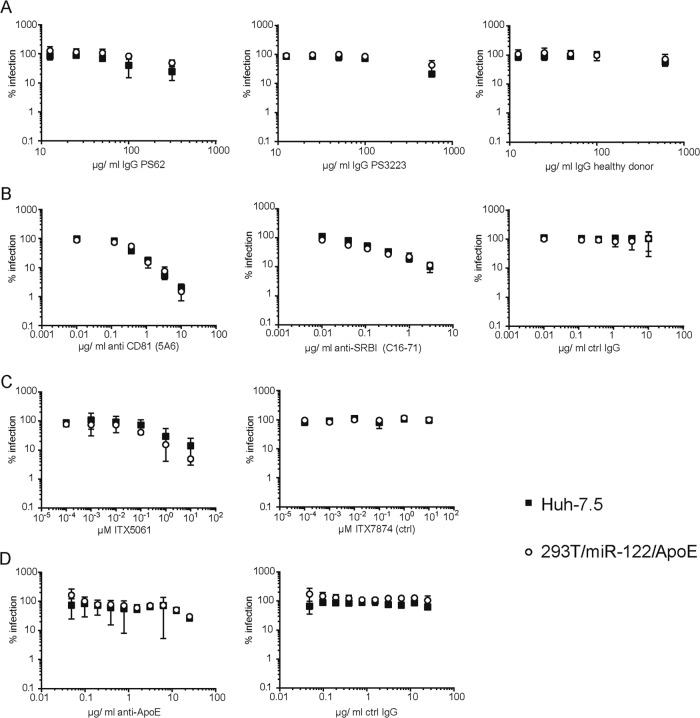
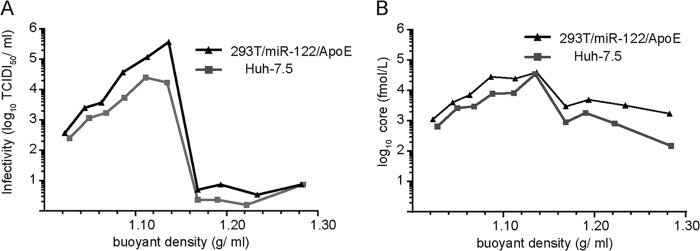
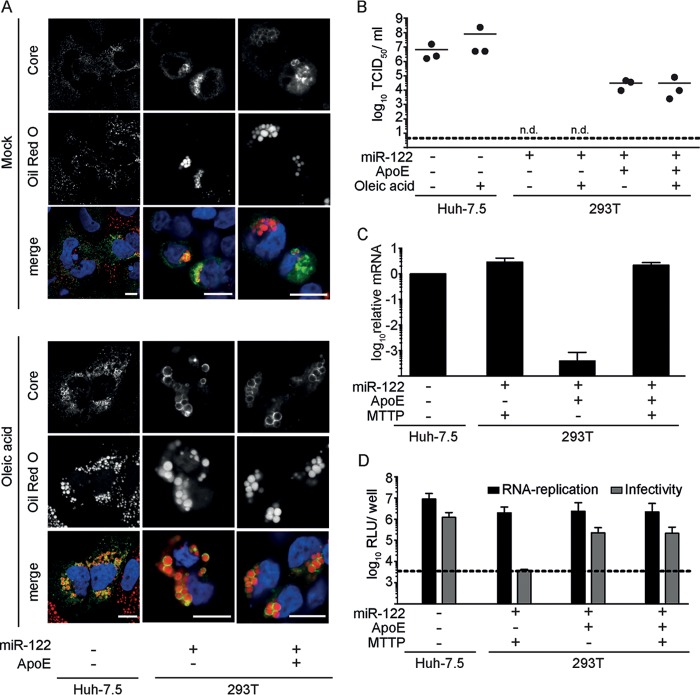
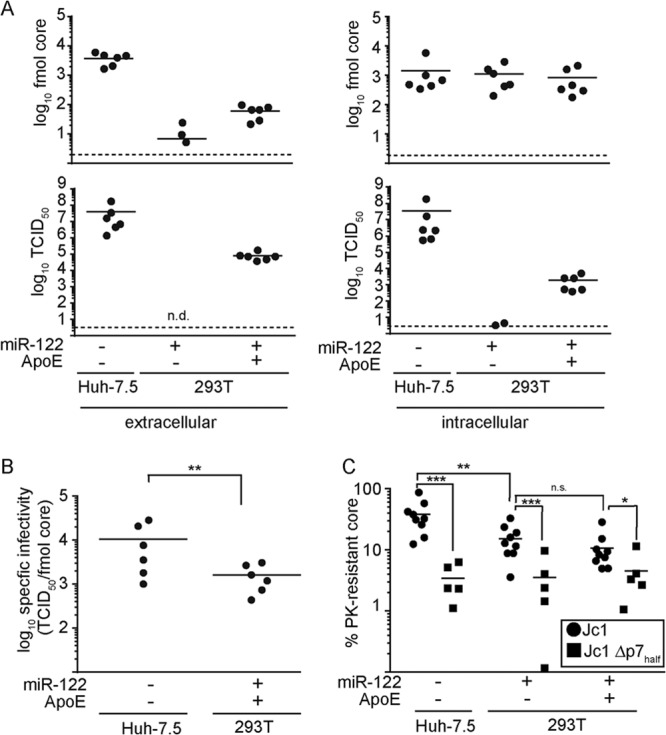
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
