

Dynamic feedback circuits function as a switch for shaping a maturation-inducing steroid pulse in Drosophila
- PMID: 24173800
- PMCID: PMC3833430
- DOI: 10.1242/dev.099739
Dynamic feedback circuits function as a switch for shaping a maturation-inducing steroid pulse in Drosophila
Abstract
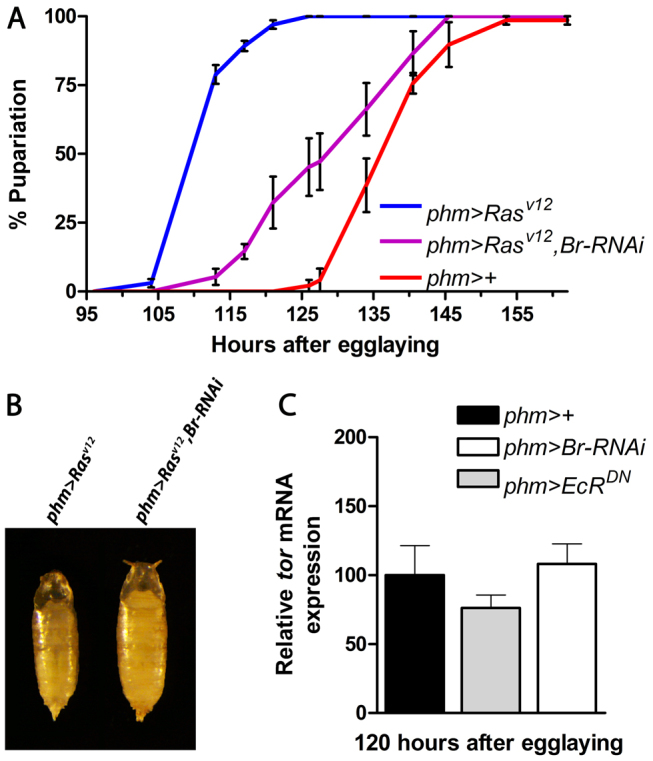
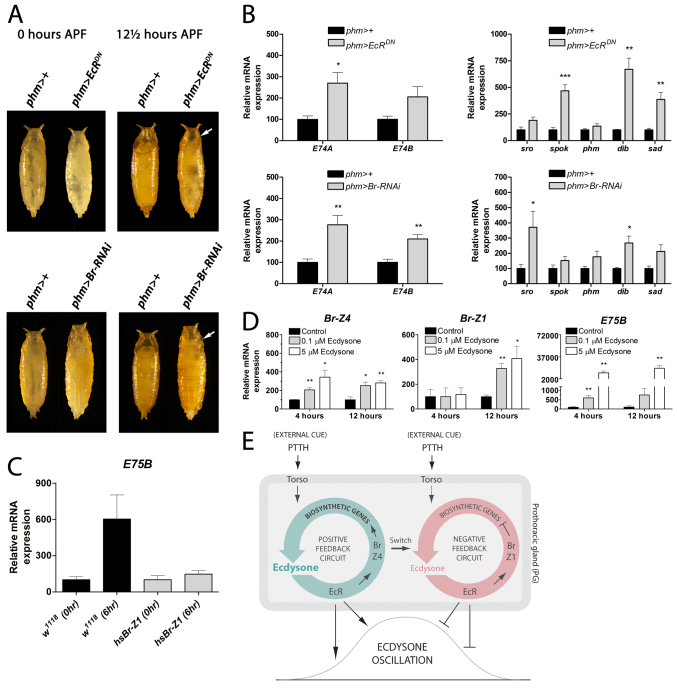
Steroid hormones trigger the onset of sexual maturation in animals by initiating genetic response programs that are determined by steroid pulse frequency, amplitude and duration. Although steroid pulses coordinate growth and timing of maturation during development, the mechanisms generating these pulses are not known. Here we show that the ecdysone steroid pulse that drives the juvenile-adult transition in Drosophila is determined by feedback circuits in the prothoracic gland (PG), the major steroid-producing tissue of insect larvae. These circuits coordinate the activation and repression of hormone synthesis, the two key parameters determining pulse shape (amplitude and duration). We show that ecdysone has a positive-feedback effect on the PG, rapidly amplifying its own synthesis to trigger pupariation as the onset of maturation. During the prepupal stage, a negative-feedback signal ensures the decline in ecdysone levels required to produce a temporal steroid pulse that drives developmental progression to adulthood. The feedback circuits rely on a developmental switch in the expression of Broad isoforms that transcriptionally activate or silence components in the ecdysone biosynthetic pathway. Remarkably, our study shows that the same well-defined genetic program that stimulates a systemic downstream response to ecdysone is also utilized upstream to set the duration and amplitude of the ecdysone pulse. Activation of this switch-like mechanism ensures a rapid, self-limiting PG response that functions in producing steroid oscillations that can guide the decision to terminate growth and promote maturation.
Keywords: Developmental timing; Ecdysone; Neuropeptide; Prothoracic gland; Prothoracicotropic hormone.
Figures
Similar articles
-
Ecdysone-dependent feedback regulation of prothoracicotropic hormone controls the timing of developmental maturation.Development. 2020 Jul 24;147(14):dev188110. doi: 10.1242/dev.188110. Development. 2020. PMID: 32631830 Free PMC article.
-
Nuclear receptor DHR4 controls the timing of steroid hormone pulses during Drosophila development.PLoS Biol. 2011 Sep;9(9):e1001160. doi: 10.1371/journal.pbio.1001160. Epub 2011 Sep 27. PLoS Biol. 2011. PMID: 21980261 Free PMC article.
-
Transcriptional control of steroid biosynthesis genes in the Drosophila prothoracic gland by ventral veins lacking and knirps.PLoS Genet. 2014 Jun 19;10(6):e1004343. doi: 10.1371/journal.pgen.1004343. eCollection 2014 Jun. PLoS Genet. 2014. PMID: 24945799 Free PMC article.
-
What goes up must come down: transcription factors have their say in making ecdysone pulses.Curr Top Dev Biol. 2013;103:35-71. doi: 10.1016/B978-0-12-385979-2.00002-2. Curr Top Dev Biol. 2013. PMID: 23347515 Review.
-
Developmental checkpoints and feedback circuits time insect maturation.Curr Top Dev Biol. 2013;103:1-33. doi: 10.1016/B978-0-12-385979-2.00001-0. Curr Top Dev Biol. 2013. PMID: 23347514 Free PMC article. Review.
Cited by
-
Endoreplication in the Drosophila melanogaster prothoracic gland is dispensable for the critical weight checkpoint.MicroPubl Biol. 2023 Feb 21;2023:10.17912/micropub.biology.000741. doi: 10.17912/micropub.biology.000741. eCollection 2023. MicroPubl Biol. 2023. PMID: 36908310 Free PMC article.
-
Expression profiling of winged- and wingless-destined pea aphid embryos implicates insulin/insulin growth factor signaling in morph differences.Evol Dev. 2020 May;22(3):257-268. doi: 10.1111/ede.12326. Epub 2019 Nov 4. Evol Dev. 2020. PMID: 31682317 Free PMC article.
-
Combgap Promotes Ovarian Niche Development and Chromatin Association of EcR-Binding Regions in BR-C.PLoS Genet. 2016 Nov 15;12(11):e1006330. doi: 10.1371/journal.pgen.1006330. eCollection 2016 Nov. PLoS Genet. 2016. PMID: 27846223 Free PMC article.
-
Timing the Juvenile-Adult Neurohormonal Transition: Functions and Evolution.Front Endocrinol (Lausanne). 2021 Feb 12;11:602285. doi: 10.3389/fendo.2020.602285. eCollection 2020. Front Endocrinol (Lausanne). 2021. PMID: 33643219 Free PMC article. Review.
-
Transcriptional regulation of insect steroid hormone biosynthesis and its role in controlling timing of molting and metamorphosis.Dev Growth Differ. 2016 Jan;58(1):94-105. doi: 10.1111/dgd.12248. Epub 2015 Dec 15. Dev Growth Differ. 2016. PMID: 26667894 Free PMC article. Review.
References
-
- Andres A. J., Fletcher J. C., Karim F. D., Thummel C. S. (1993). Molecular analysis of the initiation of insect metamorphosis: a comparative study of Drosophila ecdysteroid-regulated transcription. Dev. Biol. 160, 388–404 - PubMed
-
- Ashburner M., Chihara C., Meltzer P., Richards G. (1974). Temporal control of puffing activity in polytene chromosomes. Cold Spring Harb. Symp. Quant. Biol. 38, 655–662 - PubMed
-
- Barolo S., Carver L. A., Posakony J. W. (2000). GFP and beta-galactosidase transformation vectors for promoter/enhancer analysis in Drosophila. Biotechniques 29, 726, 728, 730, 732 - PubMed
-
- Bayer C. A., Holley B., Fristrom J. W. (1996). A switch in broad-complex zinc-finger isoform expression is regulated posttranscriptionally during the metamorphosis of Drosophila imaginal discs. Dev. Biol. 177, 1–14 - PubMed
-
- Beydon P., Lafont R. (1983). Feedback inhibition of ecdysone production by 20-hydroxyecdysone in Pieris brassicae pupae. J. Insect Physiol. 29, 529–533
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
