

Crystal structure of an HD-GYP domain cyclic-di-GMP phosphodiesterase reveals an enzyme with a novel trinuclear catalytic iron centre
- PMID: 24176013
- PMCID: PMC4159591
- DOI: 10.1111/mmi.12447
Crystal structure of an HD-GYP domain cyclic-di-GMP phosphodiesterase reveals an enzyme with a novel trinuclear catalytic iron centre
Abstract
Bis-(3',5') cyclic di-guanylate (c-di-GMP) is a key bacterial second messenger that is implicated in the regulation of many crucial processes that include biofilm formation, motility and virulence. Cellular levels of c-di-GMP are controlled through synthesis by GGDEF domain diguanylate cyclases and degradation by two classes of phosphodiesterase with EAL or HD-GYP domains. Here, we have determined the structure of an enzymatically active HD-GYP domain protein from Persephonella marina (PmGH) alone, in complex with substrate (c-di-GMP) and final reaction product (GMP). The structures reveal a novel trinuclear iron binding site, which is implicated in catalysis and identify residues involved in recognition of c-di-GMP. This structure completes the picture of all domains involved in c-di-GMP metabolism and reveals that the HD-GYP family splits into two distinct subgroups containing bi- and trinuclear metal centres.
© 2013 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Figures
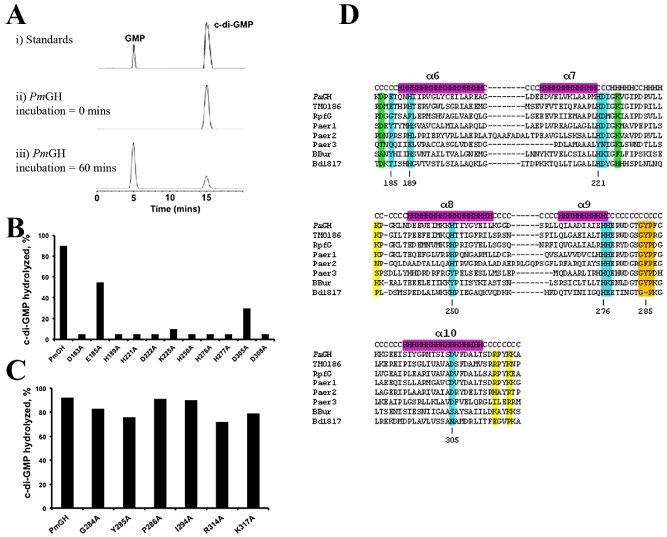
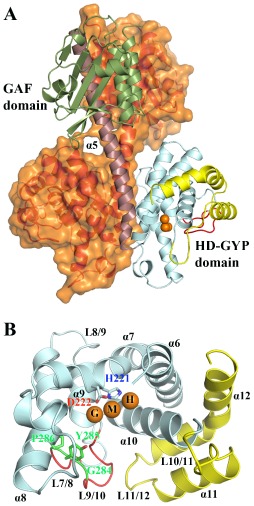
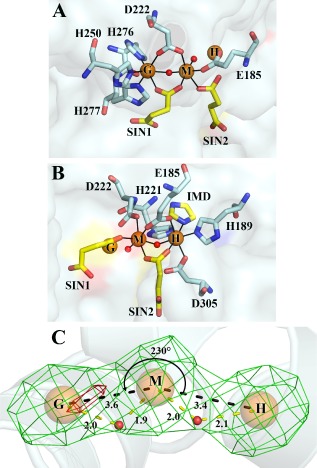
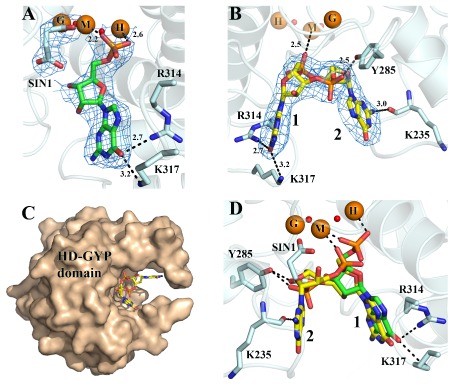

Comment in
-
Finally! The structural secrets of a HD-GYP phosphodiesterase revealed.Mol Microbiol. 2014 Jan;91(1):1-5. doi: 10.1111/mmi.12463. Epub 2013 Dec 1. Mol Microbiol. 2014. PMID: 24236493
References
-
- Aravind, L., and Koonin, E.V. (1998) The HD domain defines a new superfamily of metal‐dependent phosphohydrolases. Trends Biochem Sci 23: 469–472 - PubMed
-
- Barends, T.R., Hartmann, E., Griese, J.J., Beitlich, T., Kirienko, N.V., Ryjenkov, D.A., et al (2009) Structure and mechanism of a bacterial light‐regulated cyclic nucleotide phosphodiesterase. Nature 459: 1015–1018 - PubMed
-
- Bellini, D., Fordham‐Skelton, A., and Papiz, M.Z. (2011) STRU‐cloning: a fast, inexpensive and efficient cloning procedure applicable to both small scale and structural genomics size cloning. Mol Biotechnol 48: 30–37 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
