

PGBD5: a neural-specific intron-containing piggyBac transposase domesticated over 500 million years ago and conserved from cephalochordates to humans
- PMID: 24180413
- PMCID: PMC3902484
- DOI: 10.1186/1759-8753-4-23
PGBD5: a neural-specific intron-containing piggyBac transposase domesticated over 500 million years ago and conserved from cephalochordates to humans
Abstract
Background: piggyBac domain (PGBD) transposons are found in organisms ranging from fungi to humans. Three domesticated piggyBac elements have been described. In the ciliates Paramecium tetraurelia and Tetrahymena thermophila, homologs known as piggyMacs excise internal eliminated sequences from germline micronuclear DNA during regeneration of the new somatic macronucleus. In primates, a PGBD3 element inserted into the Cockayne syndrome group B (CSB) gene over 43 Mya serves as an alternative 3' terminal exon, enabling the CSB gene to generate both full length CSB and a conserved CSB-PGBD3 fusion protein that joins an N-terminal CSB domain to the C-terminal transposase domain.
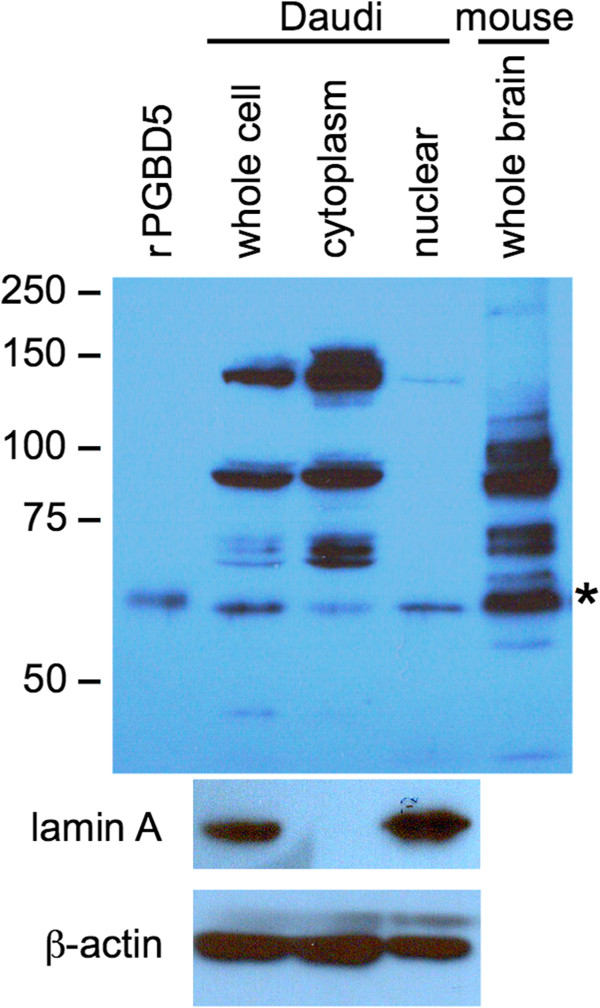
Results: We describe a fourth domesticated piggyBac element called PGBD5. We show that i) PGBD5 was first domesticated in the common ancestor of the cephalochordate Branchiostoma floridae (aka lancelet or amphioxus) and vertebrates, and is conserved in all vertebrates including lamprey but cannot be found in more basal urochordates, hemichordates, or echinoderms; ii) the lancelet, lamprey, and human PGBD5 genes are syntenic and orthologous; iii) no potentially mobile ancestral PGBD5 elements can be identified in other more deeply rooted organisms; iv) although derived from an IS4-related transposase of the RNase H clan, PGBD5 protein is unlikely to retain enzymatic activity because the catalytic DDD(D) motif is not conserved; v) PGBD5 is preferentially expressed in certain granule cell lineages of the brain and in the central nervous system based on available mouse and human in situ hybridization data, and the tissue-specificity of documented mammalian EST and mRNA clones; vi) the human PGBD5 promoter and gene region is rich in bound regulatory factors including the neuron-restrictive silencer factors NRSF/REST and CoREST, as well as SIN3, KAP1, STAT3, and CTCF; and vii) despite preferential localization within the nucleus, PGBD5 protein is unlikely to bind DNA or chromatin as neither DNase I digestion nor high salt extraction release PGBD5 from fractionated mouse brain nuclei.
Conclusions: We speculate that the neural-specific PGBD5 transposase was domesticated >500 My after cephalochordates and vertebrates split from urochordates, and that PGBD5 may have played a role in the evolution of a primitive deuterostome neural network into a centralized nervous system.
Figures









Similar articles
-
Tethering of the conserved piggyBac transposase fusion protein CSB-PGBD3 to chromosomal AP-1 proteins regulates expression of nearby genes in humans.PLoS Genet. 2012 Sep;8(9):e1002972. doi: 10.1371/journal.pgen.1002972. Epub 2012 Sep 27. PLoS Genet. 2012. PMID: 23028371 Free PMC article.
-
An abundant evolutionarily conserved CSB-PiggyBac fusion protein expressed in Cockayne syndrome.PLoS Genet. 2008 Mar 21;4(3):e1000031. doi: 10.1371/journal.pgen.1000031. PLoS Genet. 2008. PMID: 18369450 Free PMC article.
-
What role (if any) does the highly conserved CSB-PGBD3 fusion protein play in Cockayne syndrome?Mech Ageing Dev. 2013 May-Jun;134(5-6):225-33. doi: 10.1016/j.mad.2013.01.001. Epub 2013 Jan 28. Mech Ageing Dev. 2013. PMID: 23369858 Free PMC article. Review.
-
The piggyBac-derived protein 5 (PGBD5) transposes both the closely and the distantly related piggyBac-like elements Tcr-pble and Ifp2.J Mol Biol. 2021 Apr 2;433(7):166839. doi: 10.1016/j.jmb.2021.166839. Epub 2021 Feb 2. J Mol Biol. 2021. PMID: 33539889 Free PMC article.
-
Transposon domestication versus mutualism in ciliate genome rearrangements.PLoS Genet. 2013;9(8):e1003659. doi: 10.1371/journal.pgen.1003659. Epub 2013 Aug 1. PLoS Genet. 2013. PMID: 23935529 Free PMC article. Review.
Cited by
-
Compared to other NHEJ factors, DNA-PK protein and RNA levels are markedly increased in all higher primates, but not in prosimians or other mammals.DNA Repair (Amst). 2024 Oct;142:103737. doi: 10.1016/j.dnarep.2024.103737. Epub 2024 Aug 8. DNA Repair (Amst). 2024. PMID: 39128395
-
Contemporary Transposon Tools: A Review and Guide through Mechanisms and Applications of Sleeping Beauty, piggyBac and Tol2 for Genome Engineering.Int J Mol Sci. 2021 May 11;22(10):5084. doi: 10.3390/ijms22105084. Int J Mol Sci. 2021. PMID: 34064900 Free PMC article. Review.
-
The Role of Transposable Elements of the Human Genome in Neuronal Function and Pathology.Int J Mol Sci. 2022 May 23;23(10):5847. doi: 10.3390/ijms23105847. Int J Mol Sci. 2022. PMID: 35628657 Free PMC article. Review.
-
PGBD5 promotes site-specific oncogenic mutations in human tumors.Nat Genet. 2017 Jul;49(7):1005-1014. doi: 10.1038/ng.3866. Epub 2017 May 15. Nat Genet. 2017. PMID: 28504702 Free PMC article.
-
Childhood cancer mutagenesis caused by transposase-derived PGBD5.Sci Adv. 2024 Mar 22;10(12):eadn4649. doi: 10.1126/sciadv.adn4649. Epub 2024 Mar 22. Sci Adv. 2024. PMID: 38517960 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
