

Homeostatic signaling and the stabilization of neural function
- PMID: 24183022
- PMCID: PMC3856728
- DOI: 10.1016/j.neuron.2013.09.044
Homeostatic signaling and the stabilization of neural function
Abstract
The brain is astonishing in its complexity and capacity for change. This has fascinated scientists for more than a century, filling the pages of this journal for the past 25 years. But a paradigm shift is underway. It seems likely that the plasticity that drives our ability to learn and remember can only be meaningful in the context of otherwise stable, reproducible, and predictable baseline neural function. Without the existence of potent mechanisms that stabilize neural function, our capacity to learn and remember would be lost in the chaos of daily experiential change. This underscores two great mysteries in neuroscience. How are the functional properties of individual neurons and neural circuits stably maintained throughout life? And, in the face of potent stabilizing mechanisms, how can neural circuitry be modified during neural development, learning, and memory? Answers are emerging in the rapidly developing field of homeostatic plasticity.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
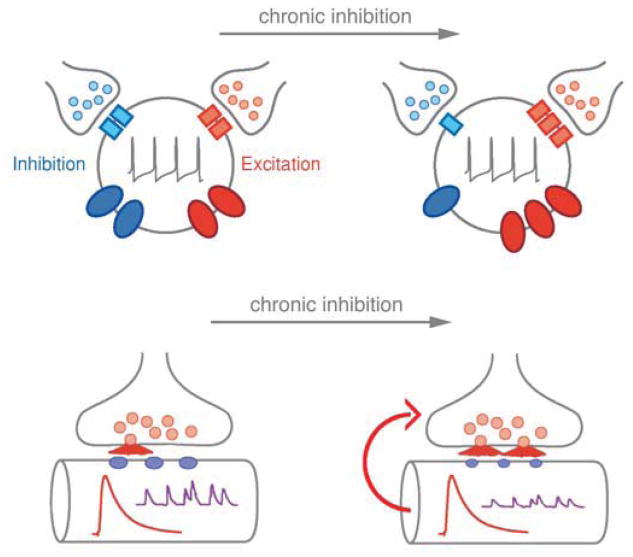
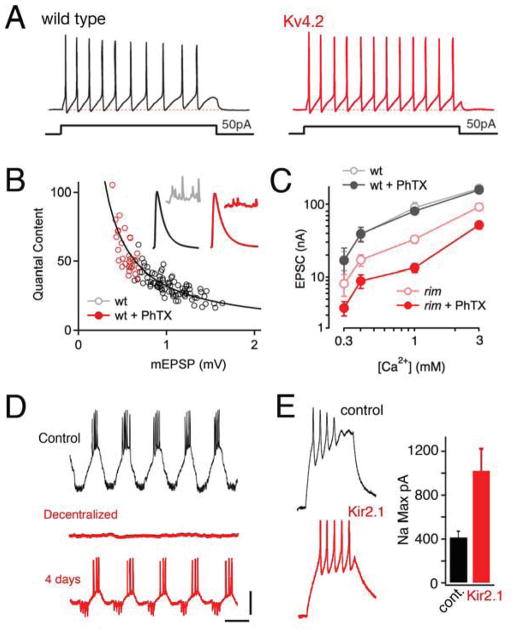
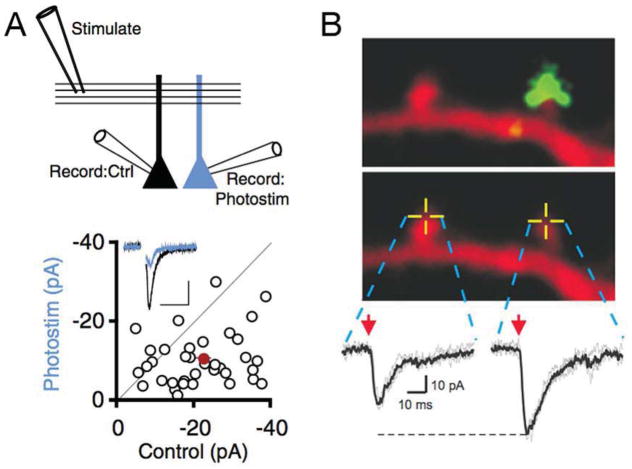
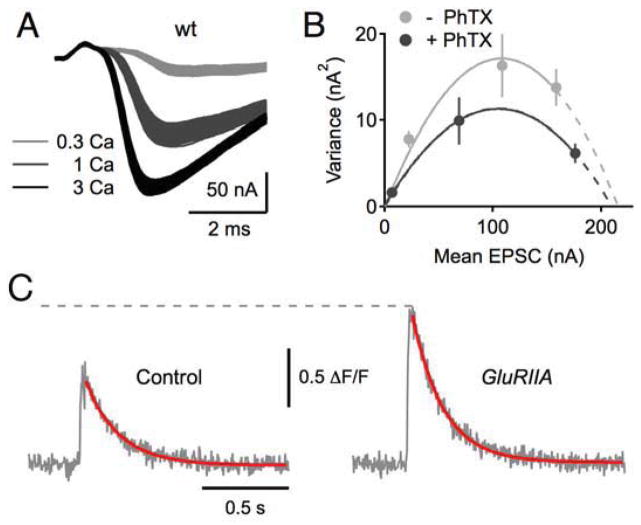
Similar articles
-
Homeostatic plasticity in neural development.Neural Dev. 2018 Jun 1;13(1):9. doi: 10.1186/s13064-018-0105-x. Neural Dev. 2018. PMID: 29855353 Free PMC article. Review.
-
Effects of cellular homeostatic intrinsic plasticity on dynamical and computational properties of biological recurrent neural networks.J Neurosci. 2013 Sep 18;33(38):15032-43. doi: 10.1523/JNEUROSCI.0870-13.2013. J Neurosci. 2013. PMID: 24048833 Free PMC article.
-
Synaptic plasticity in neural networks needs homeostasis with a fast rate detector.PLoS Comput Biol. 2013;9(11):e1003330. doi: 10.1371/journal.pcbi.1003330. Epub 2013 Nov 14. PLoS Comput Biol. 2013. PMID: 24244138 Free PMC article.
-
Postsynaptic spiking homeostatically induces cell-autonomous regulation of inhibitory inputs via retrograde signaling.J Neurosci. 2010 Dec 1;30(48):16220-31. doi: 10.1523/JNEUROSCI.3085-10.2010. J Neurosci. 2010. PMID: 21123568 Free PMC article.
-
Homeostatic plasticity-a presynaptic perspective.Curr Opin Neurobiol. 2019 Feb;54:155-162. doi: 10.1016/j.conb.2018.10.003. Epub 2018 Oct 25. Curr Opin Neurobiol. 2019. PMID: 30384022 Review.
Cited by
-
Modulator-Gated, SUMOylation-Mediated, Activity-Dependent Regulation of Ionic Current Densities Contributes to Short-Term Activity Homeostasis.J Neurosci. 2019 Jan 23;39(4):596-611. doi: 10.1523/JNEUROSCI.1379-18.2018. Epub 2018 Nov 30. J Neurosci. 2019. PMID: 30504282 Free PMC article.
-
IGF-1 receptor regulates upward firing rate homeostasis via the mitochondrial calcium uniporter.Proc Natl Acad Sci U S A. 2022 Aug 16;119(33):e2121040119. doi: 10.1073/pnas.2121040119. Epub 2022 Aug 9. Proc Natl Acad Sci U S A. 2022. PMID: 35943986 Free PMC article.
-
Juvenile Social Isolation Enhances the Activity of Inhibitory Neuronal Circuits in the Medial Prefrontal Cortex.Front Cell Neurosci. 2020 May 12;14:105. doi: 10.3389/fncel.2020.00105. eCollection 2020. Front Cell Neurosci. 2020. PMID: 32477068 Free PMC article.
-
Combining HF rTMS over the Left DLPFC with Concurrent Cognitive Activity for the Offline Modulation of Working Memory in Healthy Volunteers: A Proof-of-Concept Study.Brain Sci. 2020 Feb 4;10(2):83. doi: 10.3390/brainsci10020083. Brain Sci. 2020. PMID: 32033106 Free PMC article.
-
Altered active zones, vesicle pools, nerve terminal conductivity, and morphology during experimental MuSK myasthenia gravis.PLoS One. 2014 Dec 1;9(12):e110571. doi: 10.1371/journal.pone.0110571. eCollection 2014. PLoS One. 2014. PMID: 25438154 Free PMC article.
References
-
- Awatramani GB, Price GD, Trussell LO. Modulation of Transmitter Release by Presynaptic Resting Potential and Background Calcium Levels. Neuron. 2005;48:109–121. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
