

Cerebellar development in the absence of Gbx function in zebrafish
- PMID: 24183937
- PMCID: PMC3935510
- DOI: 10.1016/j.ydbio.2013.10.026
Cerebellar development in the absence of Gbx function in zebrafish
Abstract
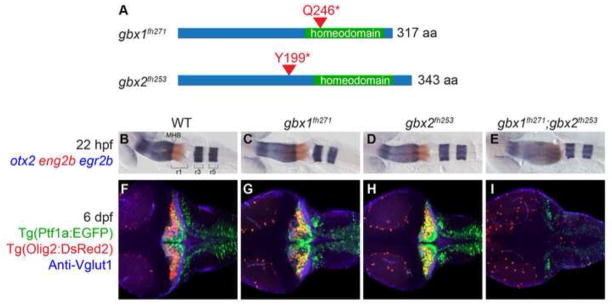
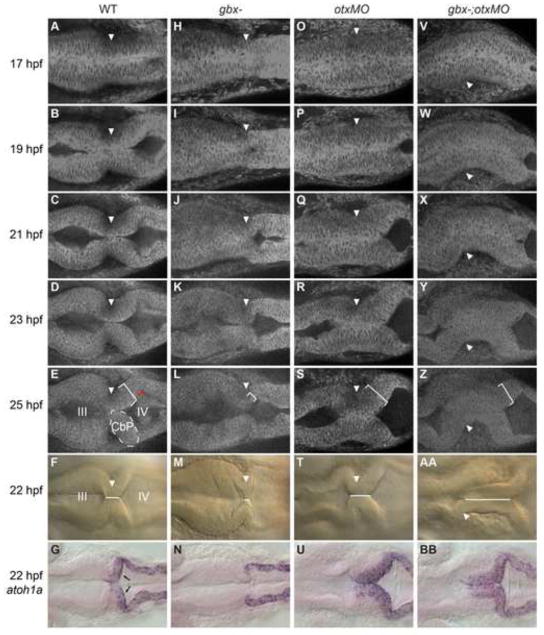
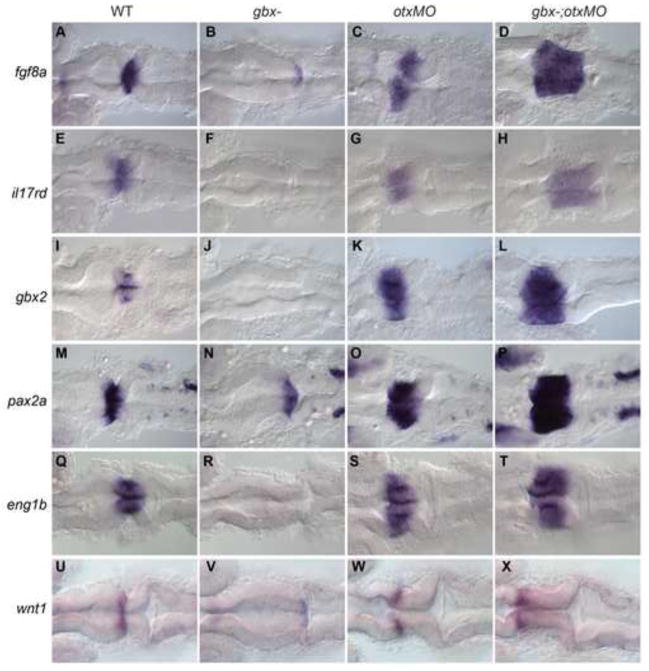
The midbrain-hindbrain boundary (MHB) is a well-known organizing center during vertebrate brain development. The MHB forms at the expression boundary of Otx2 and Gbx2, mutually repressive homeodomain transcription factors expressed in the midbrain/forebrain and anterior hindbrain, respectively. The genetic hierarchy of gene expression at the MHB is complex, involving multiple positive and negative feedback loops that result in the establishment of non-overlapping domains of Wnt1 and Fgf8 on either side of the boundary and the consequent specification of the cerebellum. The cerebellum derives from the dorsal part of the anterior-most hindbrain segment, rhombomere 1 (r1), which undergoes a distinctive morphogenesis to give rise to the cerebellar primordium within which the various cerebellar neuron types are specified. Previous studies in the mouse have shown that Gbx2 is essential for cerebellar development. Using zebrafish mutants we show here that in the zebrafish gbx1 and gbx2 are required redundantly for morphogenesis of the cerebellar primordium and subsequent cerebellar differentiation, but that this requirement is alleviated by knocking down Otx. Expression of fgf8, wnt1 and the entire MHB genetic program is progressively lost in gbx1-;gbx2- double mutants but is rescued by Otx knock-down. This rescue of the MHB genetic program depends on rescued Fgf signaling, however the rescue of cerebellar primordium morphogenesis is independent of both Gbx and Fgf. Based on our findings we propose a revised model for the role of Gbx in cerebellar development.
Keywords: Cerebellum; Gbx; Zebrafish.
© 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Acampora D, Avantaggiato V, Tuorto F, Briata P, Corte G, Simeone A. Visceral endoderm-restricted translation of Otx1 mediates recovery of Otx2 requirements for specification of anterior neural plate and normal gastrulation. Development. 1998;125:5091–104. - PubMed
-
- Bae YK, Kani S, Shimizu T, Tanabe K, Nojima H, Kimura Y, Higashijima S, Hibi M. Anatomy of zebrafish cerebellum and screen for mutations affecting its development. Dev Biol. 2009;330:406–26. - PubMed
-
- Bally-Cuif L, Cholley B, Wassef M. Involvement of Wnt-1 in the formation of the mes/metencephalic boundary. Mech Dev. 1995;53:23–34. - PubMed
-
- Broccoli V, Boncinelli E, Wurst W. The caudal limit of Otx2 expression positions the isthmic organizer. Nature. 1999;401:164–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
