

Polo-like kinase 4 autodestructs by generating its Slimb-binding phosphodegron
- PMID: 24184097
- PMCID: PMC3844517
- DOI: 10.1016/j.cub.2013.09.019
Polo-like kinase 4 autodestructs by generating its Slimb-binding phosphodegron
Abstract
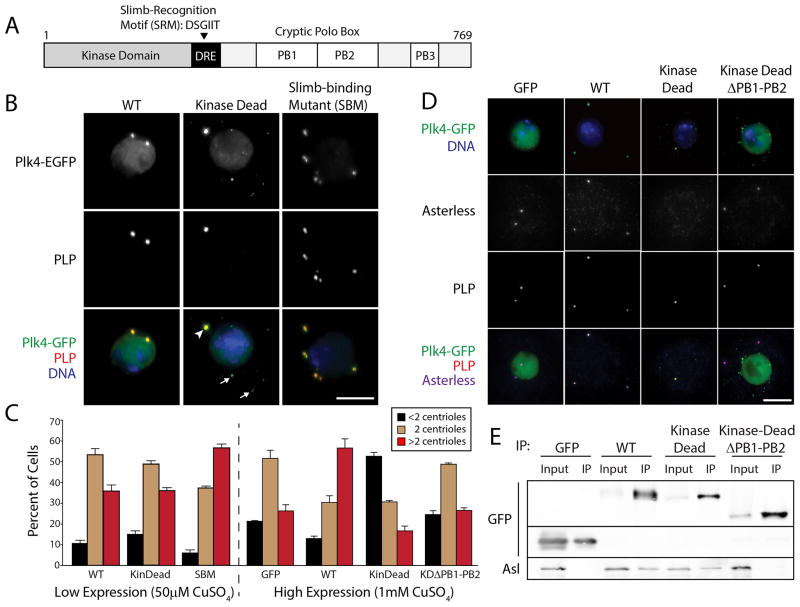
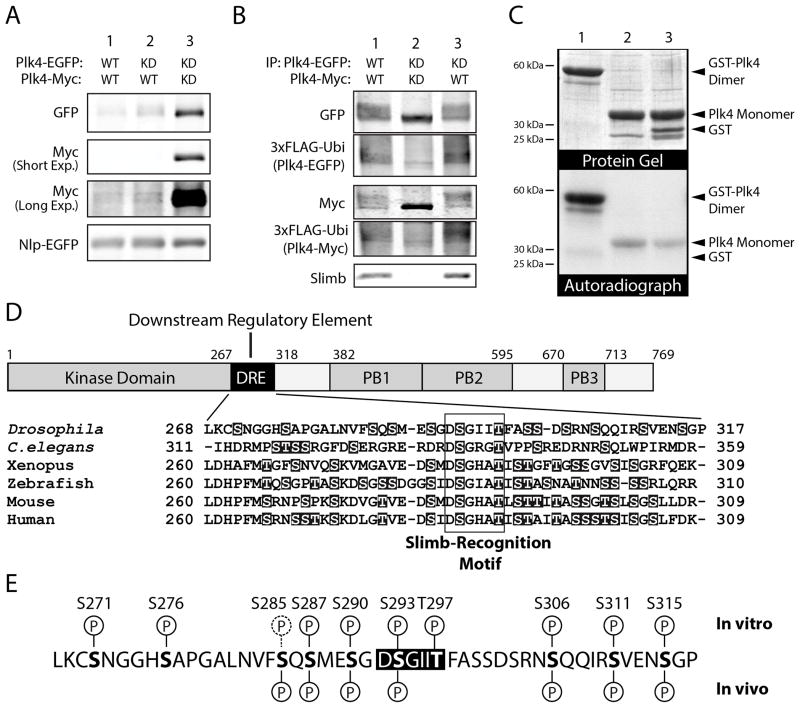
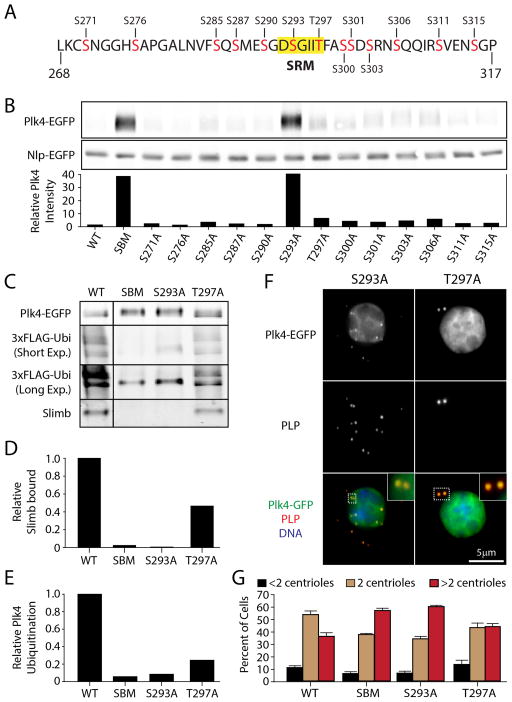
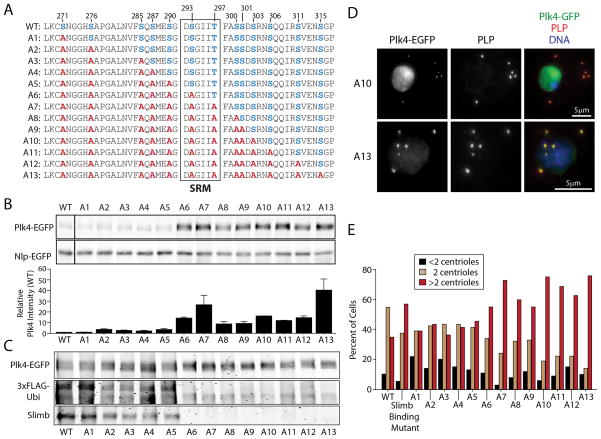
Polo-like kinase 4 (Plk4) is a conserved master regulator of centriole assembly. Previously, we found that Drosophila Plk4 protein levels are actively suppressed during interphase. Degradation of interphase Plk4 prevents centriole overduplication and is mediated by the ubiquitin-ligase complex SCF(Slimb/βTrCP). Since Plk4 stability depends on its activity, we studied the consequences of inactivating Plk4 or perturbing its phosphorylation state within its Slimb-recognition motif (SRM). Mass spectrometry of in-vitro-phosphorylated Plk4 and Plk4 purified from cells reveals that it is directly responsible for extensively autophosphorylating and generating its Slimb-binding phosphodegron. Phosphorylatable residues within this regulatory region were systematically mutated to determine their impact on Plk4 protein levels and centriole duplication when expressed in S2 cells. Notably, autophosphorylation of a single residue (Ser293) within the SRM is critical for Slimb binding and ubiquitination. Our data also demonstrate that autophosphorylation of numerous residues flanking S293 collectively contribute to establishing a high-affinity binding site for SCF(Slimb). Taken together, our findings suggest that Plk4 directly generates its own phosphodegron and can do so without the assistance of an additional kinase(s).
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Brito DA, Gouveia SM, Bettencourt-Dias M. Deconstructing the centriole: structure and number control. Curr Opin Cell Biol. 2012;24:4–13. - PubMed
-
- Cunha-Ferreira I, Rodrigues-Martins A, Bento I, Riparbeli M, Zhang W, Laue E, Callaini G, Glover DM, Bettencourt-Dias M. The SCF/Slimb ubiquitin ligase limits centrosome amplification through degradation of SAK/PLK4. Curr Biol. 2009;19:43–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
