

Polyploidization and cell fusion contribute to wound healing in the adult Drosophila epithelium
- PMID: 24184101
- PMCID: PMC3898104
- DOI: 10.1016/j.cub.2013.09.029
Polyploidization and cell fusion contribute to wound healing in the adult Drosophila epithelium
Abstract
Background: Reestablishing epithelial integrity and biosynthetic capacity is critically important following tissue damage. The adult Drosophila abdominal epithelium provides an attractive new system to address how postmitotic diploid cells contribute to repair.
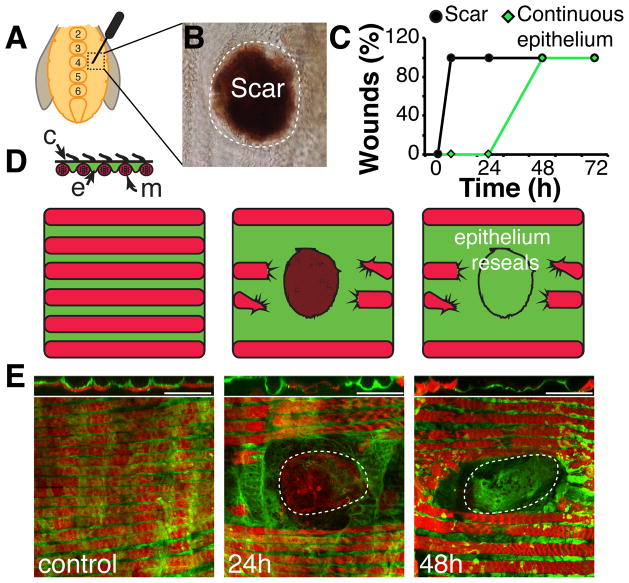
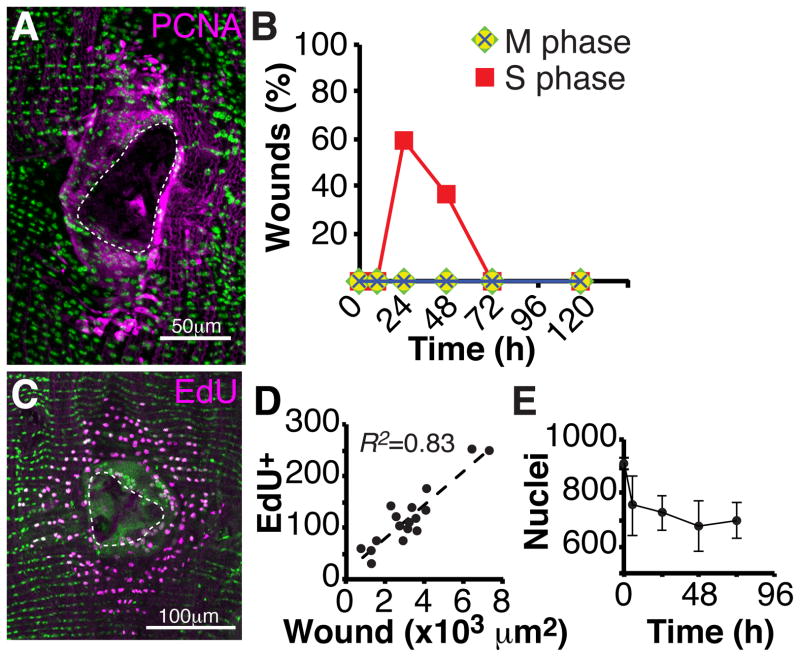
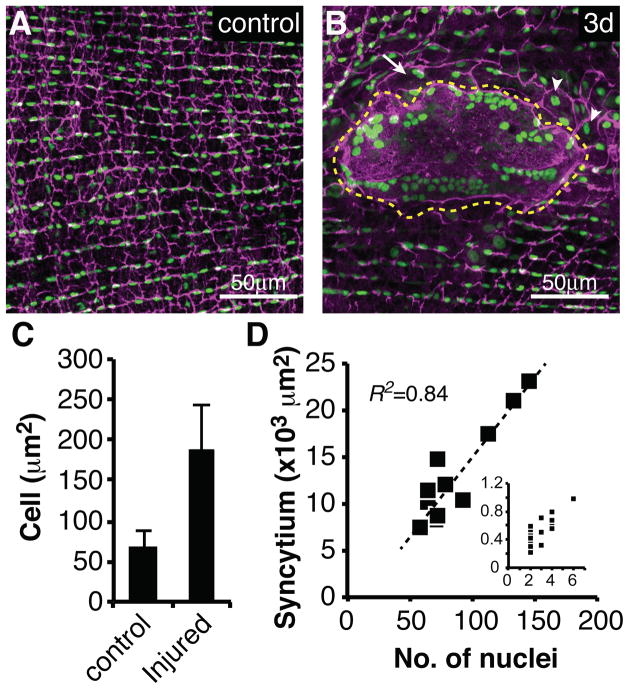
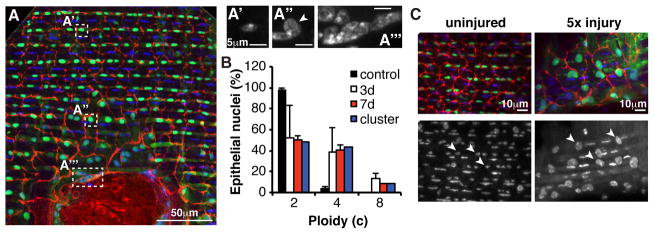
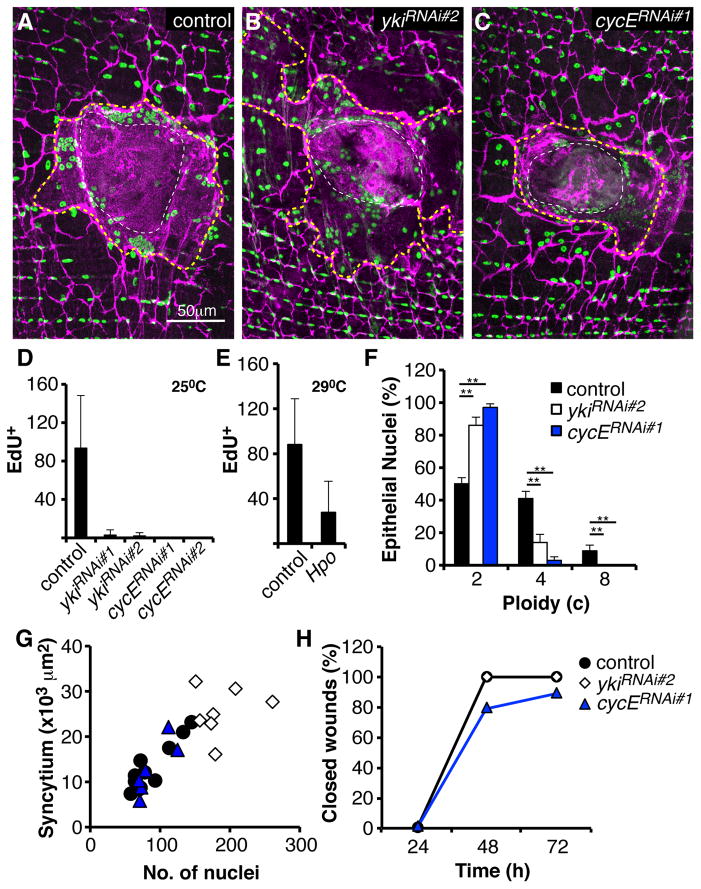
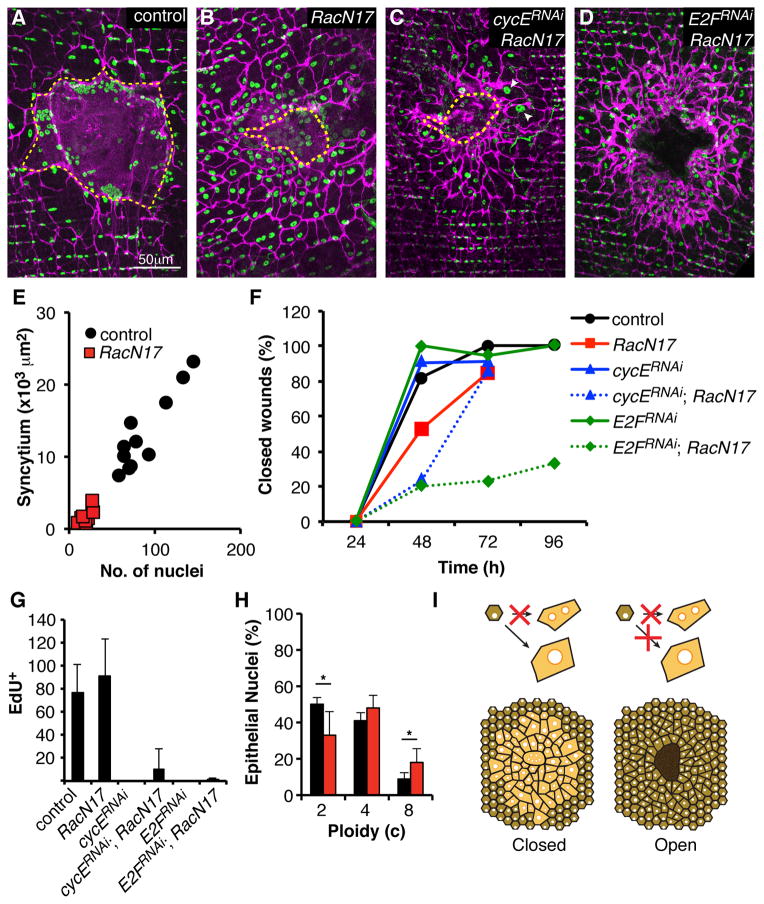
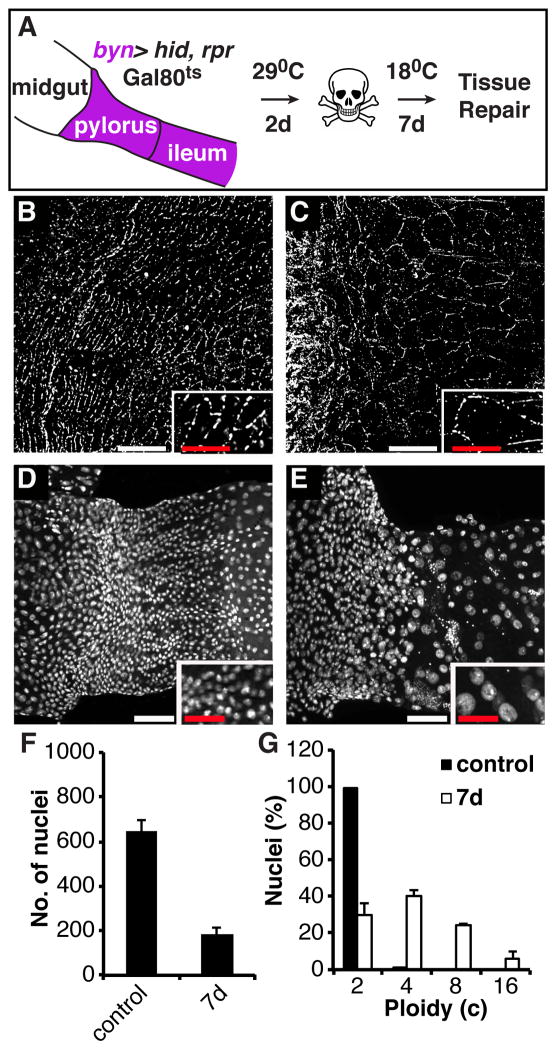
Results: Puncture wounds to the adult Drosophila epidermis close initially by forming a melanized scab. We found that epithelial cells near the wound site fuse to form a giant syncytium, which sends lamellae under the scab to re-epithelialize the damaged site. Other large cells arise more peripherally by initiating endocycles and becoming polyploid, or by cell fusion. Rac GTPase activity is needed for syncytium formation, while the Hippo signaling effector Yorkie modulates both polyploidization and cell fusion. Large cell formation is functionally important because when both polyploidization and fusion are blocked, wounds do not re-epithelialize.
Conclusions: Our observations indicate that cell mass lost upon wounding can be replaced by polyploidization instead of mitotic proliferation. We propose that large cells generated by polyploidization or cell fusion are essential because they are better able than diploid cells to mechanically stabilize wounds, especially those containing permanent acellular structures, such as scar tissue.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
