

Enhancing T lineage production in aged mice: a novel function of Foxn1 in the bone marrow niche
- PMID: 24184560
- PMCID: PMC4081451
- DOI: 10.4049/jimmunol.1202278
Enhancing T lineage production in aged mice: a novel function of Foxn1 in the bone marrow niche
Abstract
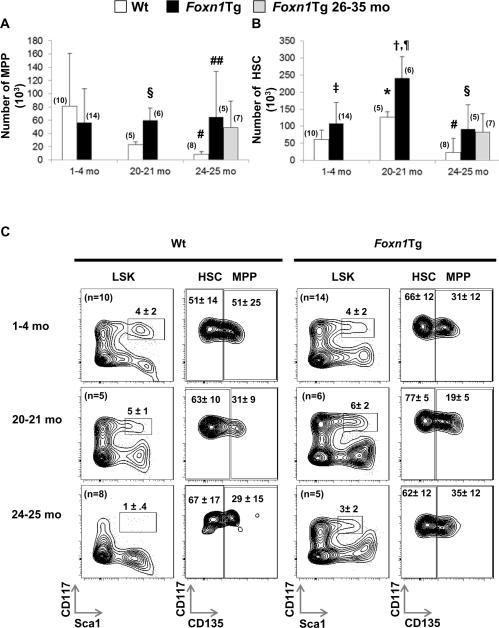
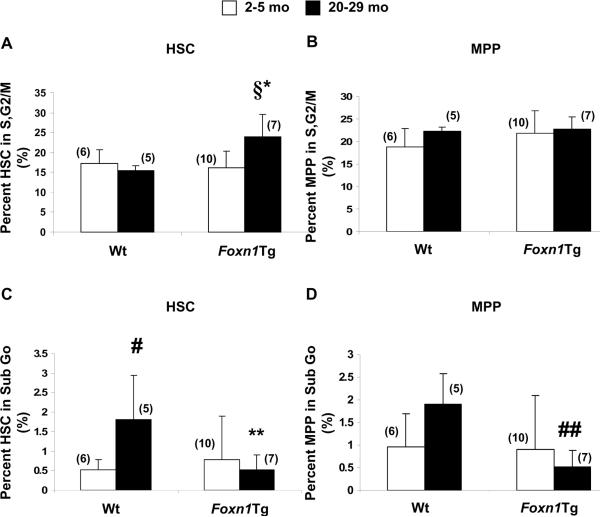
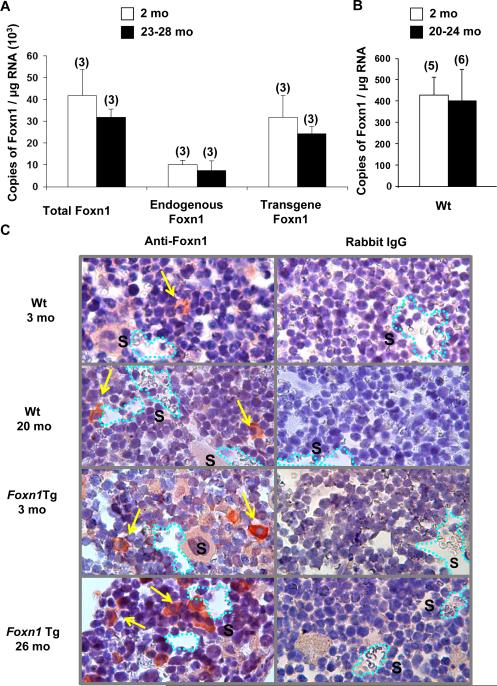
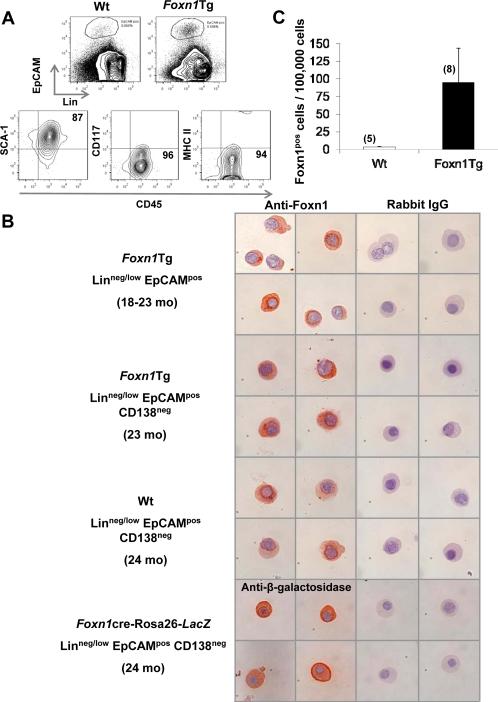
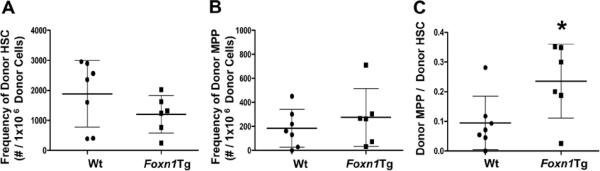
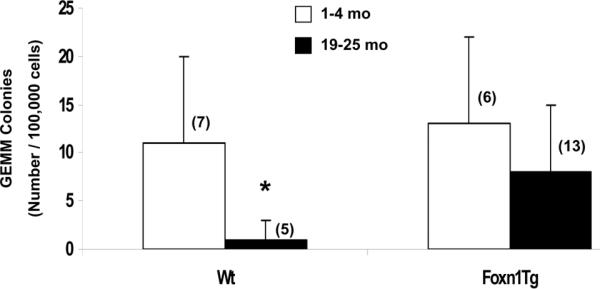
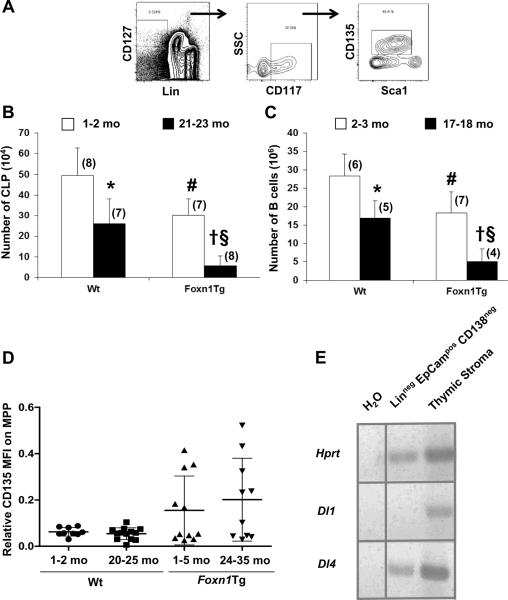
Foxn1 is essential for thymic organogenesis and T lymphopoiesis. Whereas reduced Foxn1 expression results in a decline in T lymphopoiesis, overexpression of Foxn1 in the thymus of a transgenic mouse model (Foxn1Tg) attenuates the age-associated decline in T lymphopoiesis. T lymphopoiesis begins with early T cell progenitors (ETP), derived from multipotent progenitors (MPP) in the bone marrow (BM). A decline in MPP and ETP numbers with age is thought to contribute to reduced T lymphopoiesis. Previously, we showed that reduced ETP number with age is attenuated in Foxn1 transgenic (Tg); whether the effect is initiated in the BM with MPP is not known. In this study, we report that Foxn1 is expressed in wild-type BM and overexpressed in Foxn1Tg. With age, the number of MPP in Foxn1Tg was not reduced, and Foxn1Tg also have a larger pool of hematopoietic stem cells. Furthermore, the Foxn1Tg BM is more efficient in generating MPP. In contrast to MPP, common lymphoid progenitors and B lineage cell numbers were significantly lower in both young and aged Foxn1Tg compared with wild type. We identified a novel population of lineage(neg/low), CD45(pos) EpCAM(pos), SCA1(pos), CD117(neg), CD138(neg), MHCII(neg) cells as Foxn1-expressing BM cells that also express Delta-like 4. Thus, Foxn1 affects both T lymphopoiesis and hematopoiesis, and the Foxn1 BM niche may function in skewing MPP development toward T lineage progenitors.
Figures
References
-
- Bleul CC, Corbeaux T, Reuter A, Fisch P, Monting JS, Boehm T. Formation of a functional thymus initiated by a postnatal epithelial progenitor cell. Nature. 2006;441:992–996. - PubMed
-
- Nehls M, Pfeifer D, Schorpp M, Hedrich H, Boehm T. New member of the winged-helix protein family disrupted in mouse and rat nude mutations. Nature. 1994;372:103–107. - PubMed
-
- Ortman CL, Dittmar KA, Witte PL, Le PT. Molecular characterization of the mouse involuted thymus: aberrations in expression of transcription regulators in thymocyte and epithelial compartments. Int. Immunol. 2002;14:813–822. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
