

Regulation of interneuron excitability by gap junction coupling with principal cells
- PMID: 24185427
- PMCID: PMC3963432
- DOI: 10.1038/nn.3569
Regulation of interneuron excitability by gap junction coupling with principal cells
Abstract
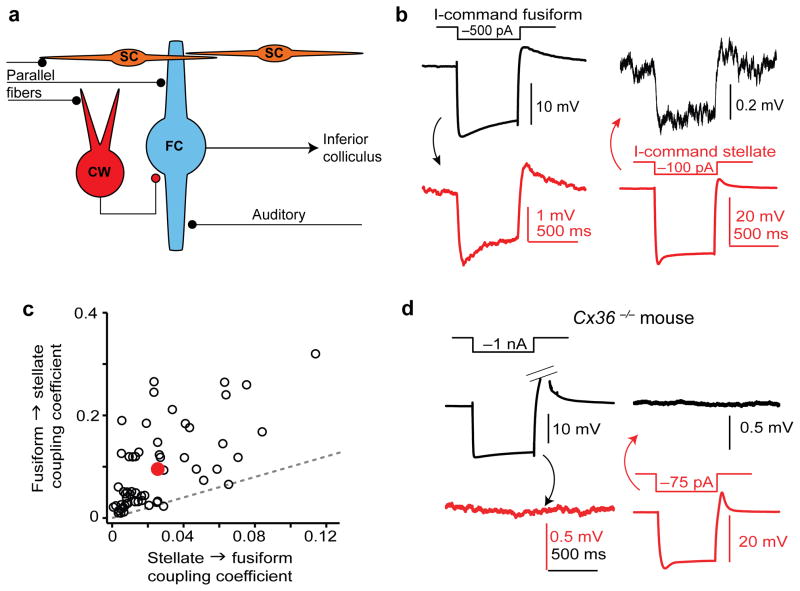
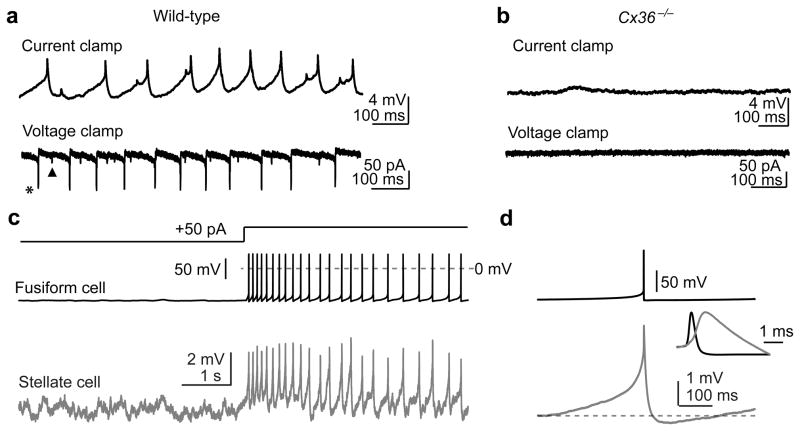
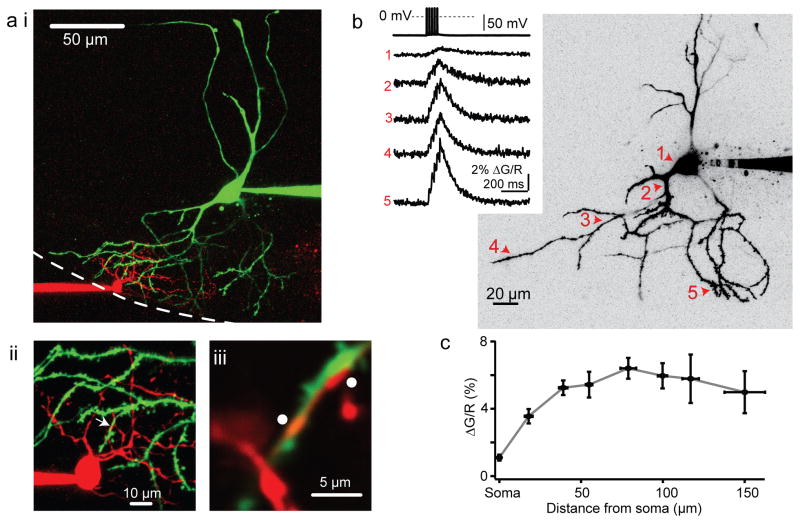
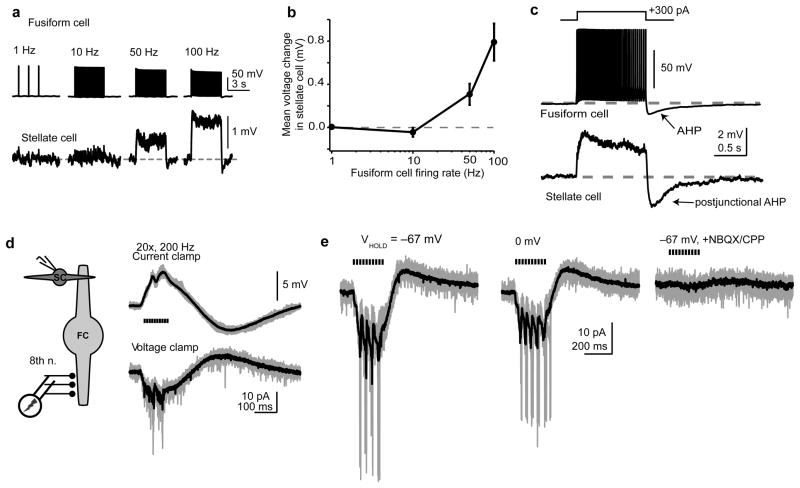
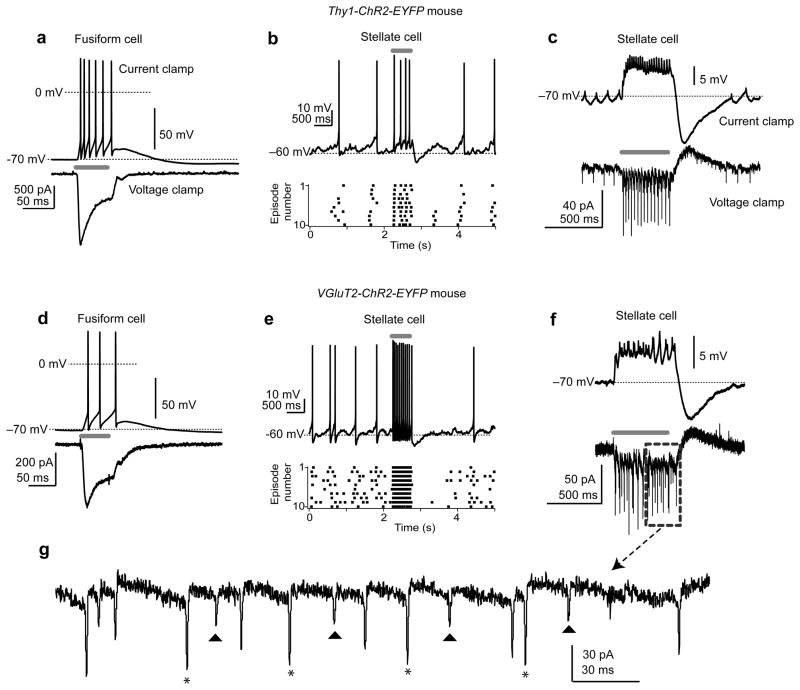
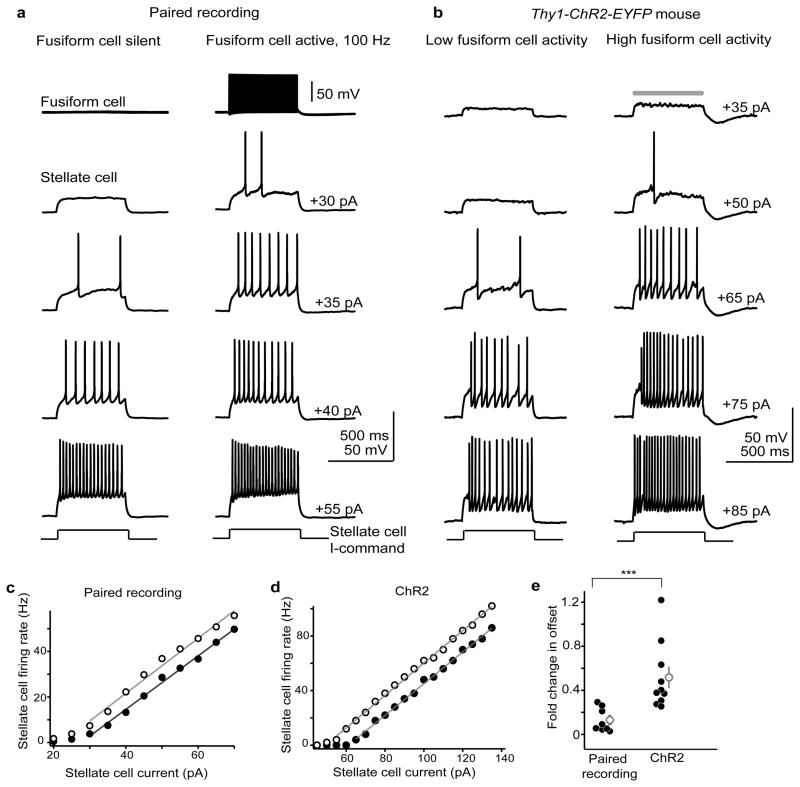
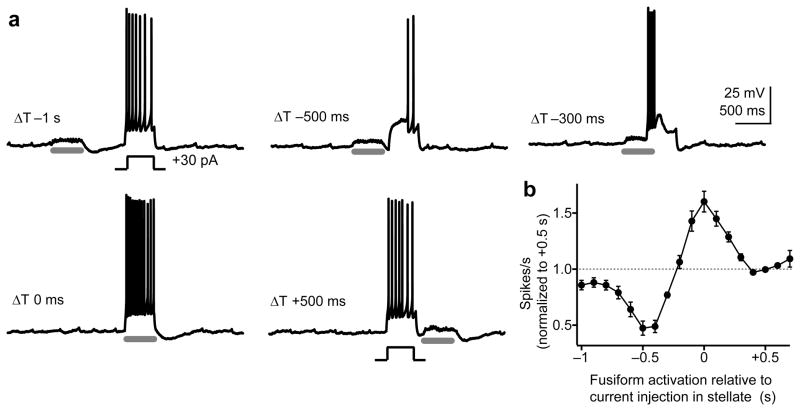
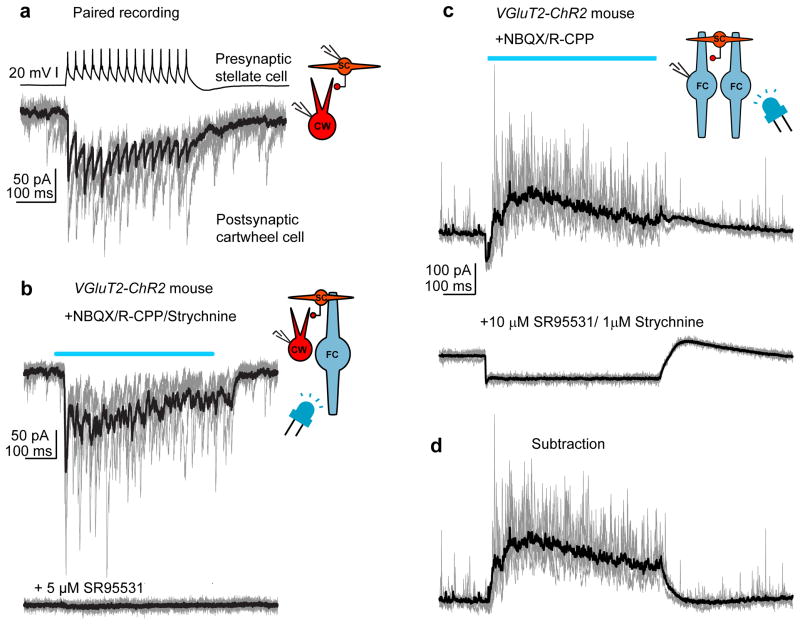
Electrical coupling of inhibitory interneurons can synchronize activity across multiple neurons, thereby enhancing the reliability of inhibition onto principal cell targets. It is unclear whether downstream activity in principal cells controls the excitability of such inhibitory networks. Using paired patch-clamp recordings, we show that excitatory projection neurons (fusiform cells) and inhibitory stellate interneurons of the dorsal cochlear nucleus form an electrically coupled network through gap junctions containing connexin36 (Cxc36, also called Gjd2). Remarkably, stellate cells were more strongly coupled to fusiform cells than to other stellate cells. This heterologous coupling was functionally asymmetric, biasing electrical transmission from the principal cell to the interneuron. Optogenetically activated populations of fusiform cells reliably enhanced interneuron excitability and generated GABAergic inhibition onto the postsynaptic targets of stellate cells, whereas deep afterhyperpolarizations following fusiform cell spike trains potently inhibited stellate cells over several hundred milliseconds. Thus, the excitability of an interneuron network is bidirectionally controlled by distinct epochs of activity in principal cells.
Figures
References
-
- Bell CC, Han V, Sawtell NB. Cerebellum-like structures and their implications for cerebellar function. Annu Rev Neurosci. 2008;31:1–24. - PubMed
-
- Dean P, Porrill J, Ekerot CF, Jörntell H. The cerebellar microcircuit as an adaptive filter: experimental and computational evidence. Nat Rev Neurosci. 2010;11:30–43. - PubMed
-
- Requarth T, Sawtell NB. Neural mechanisms for filtering self-generated sensory signals in cerebellum-like circuits. Curr Opin Neurobiol. 2011;21:602–608. - PubMed
-
- Oertel D, Young ED. What’s a cerebellar circuit doing in the auditory system? Trends Neurosci. 2004;27:104–110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
