

Development of functionally selective, small molecule agonists at kappa opioid receptors
- PMID: 24187130
- PMCID: PMC3868780
- DOI: 10.1074/jbc.M113.504381
Development of functionally selective, small molecule agonists at kappa opioid receptors
Abstract
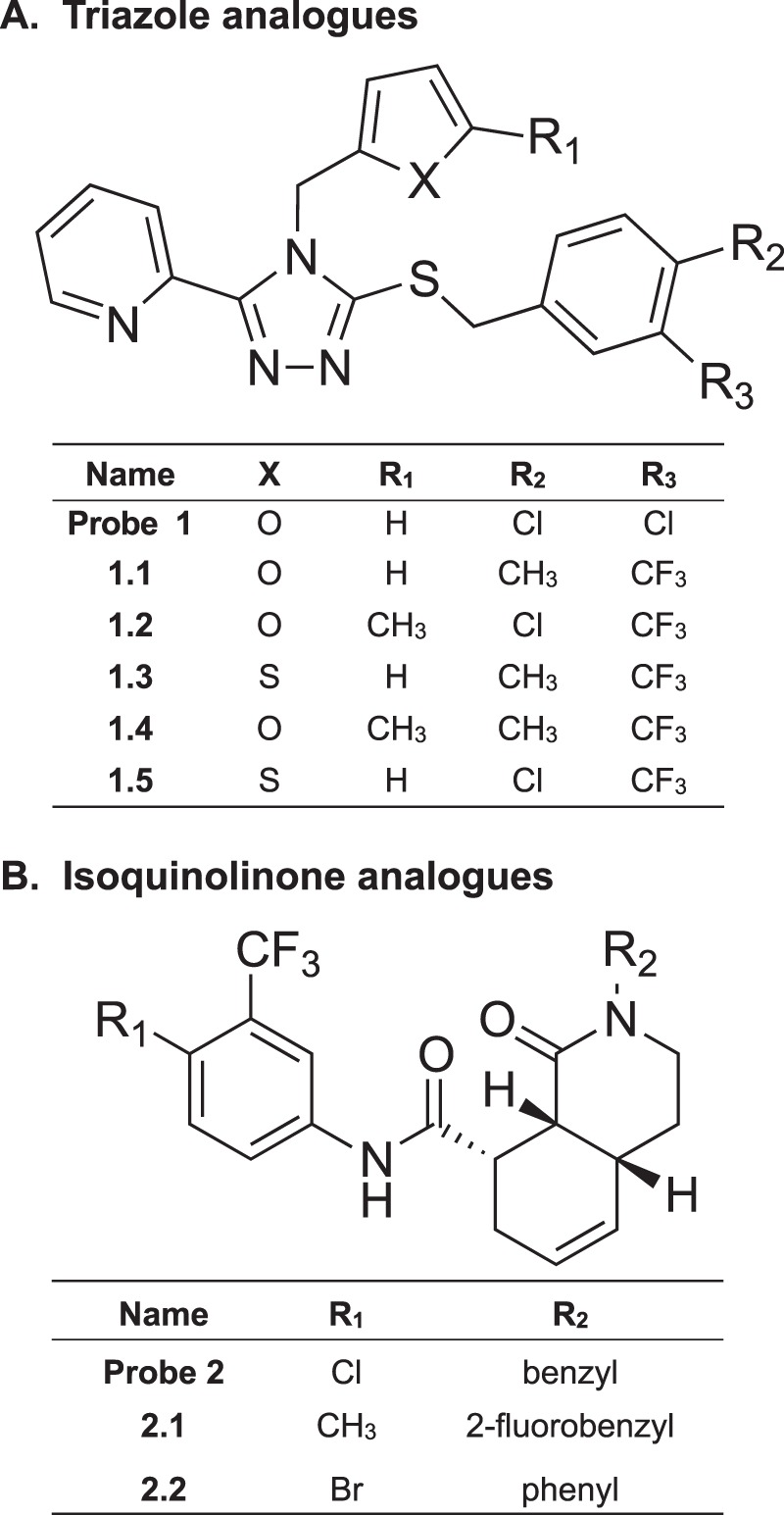
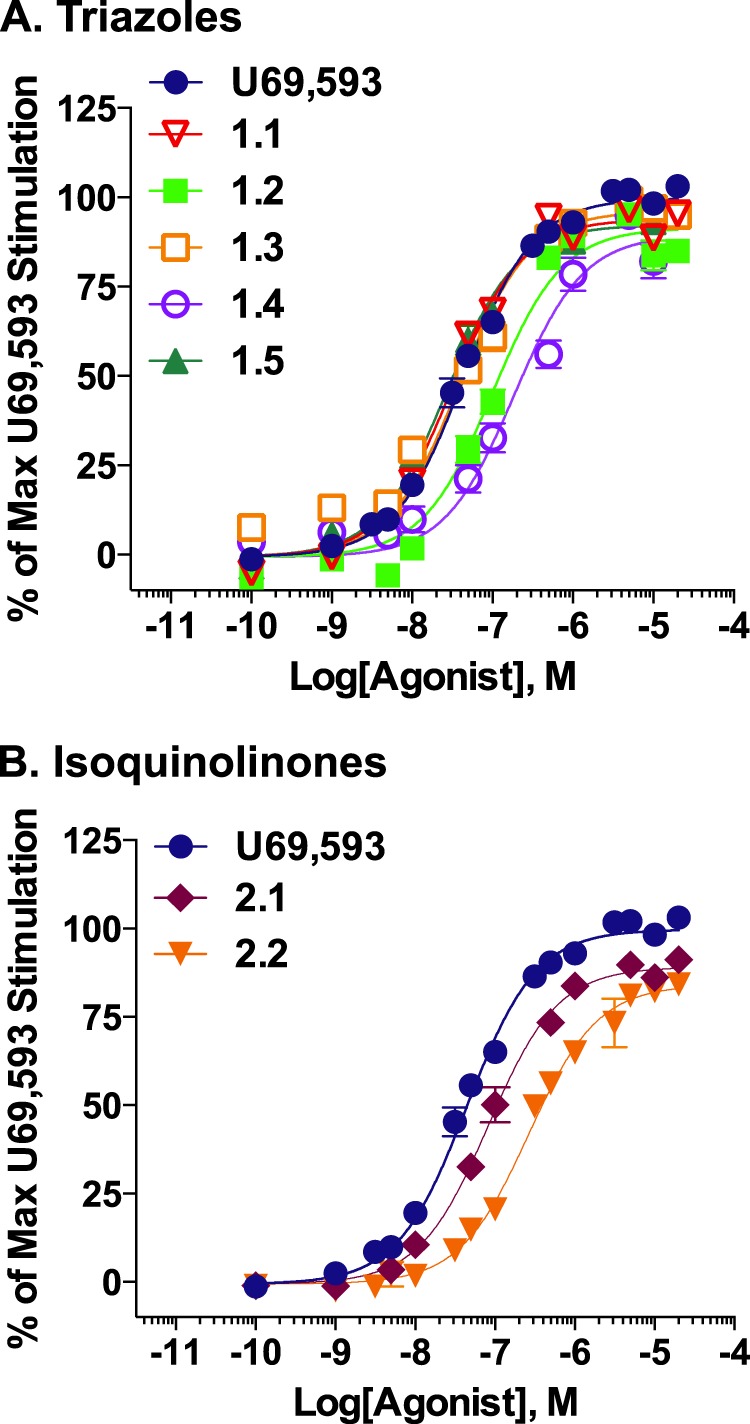
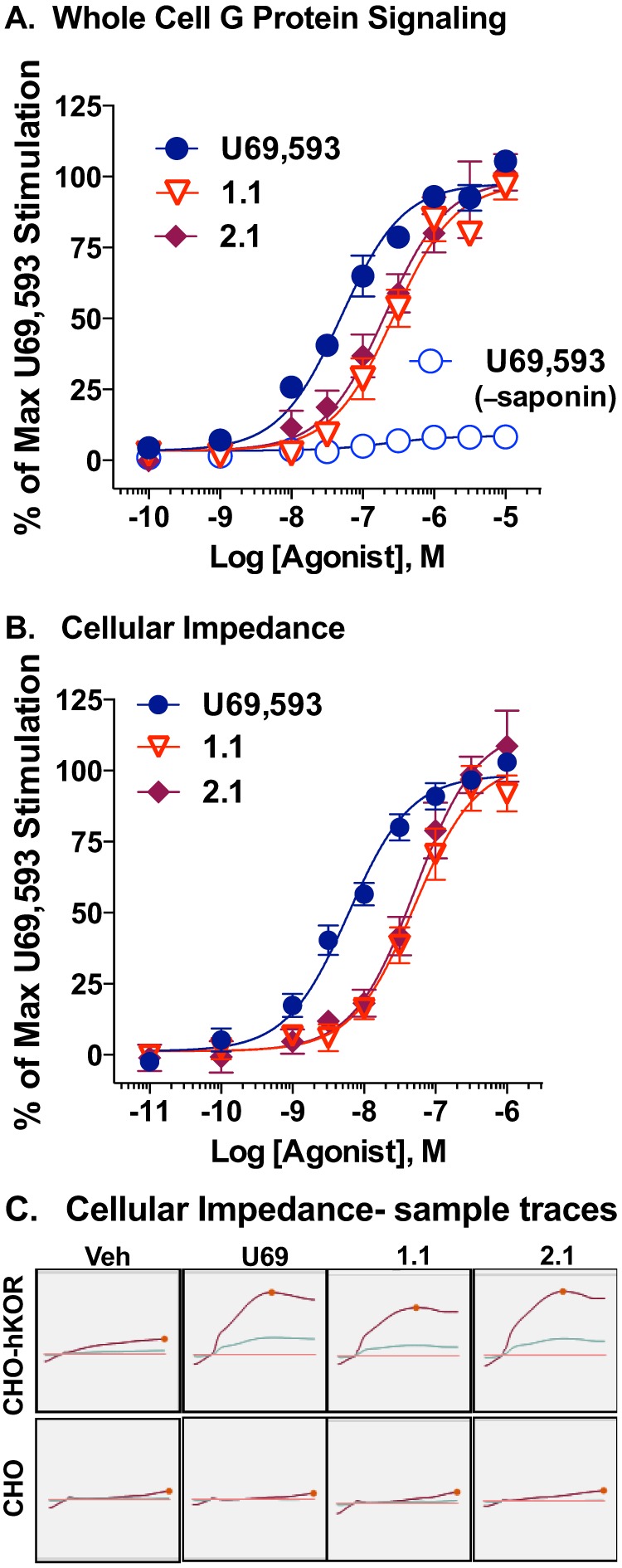
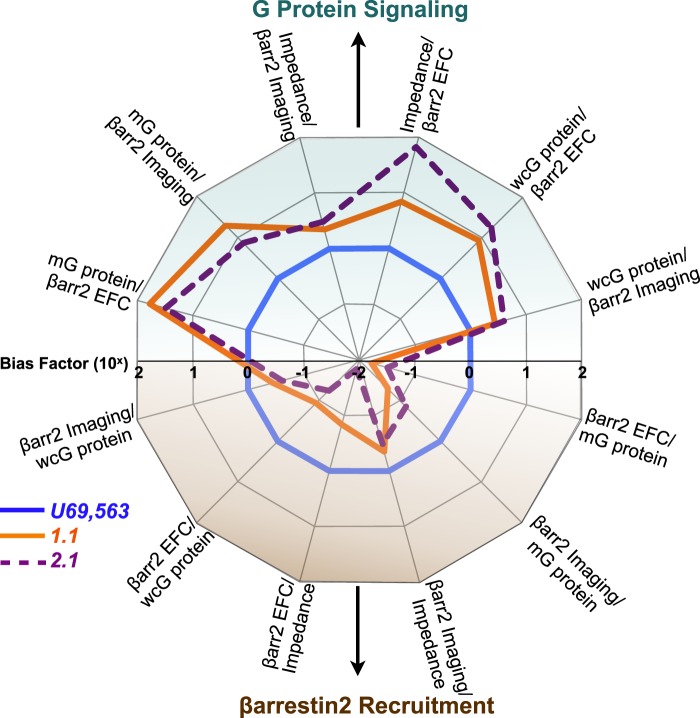
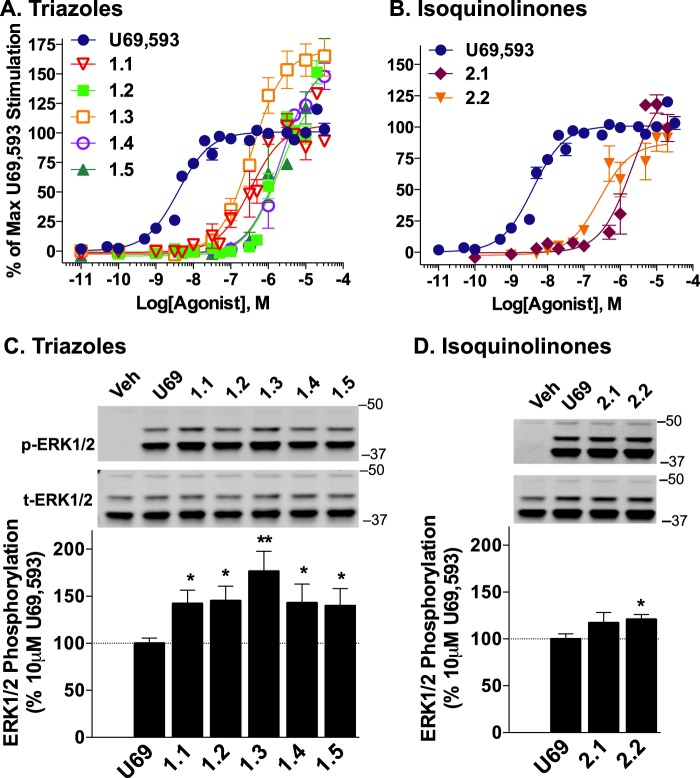
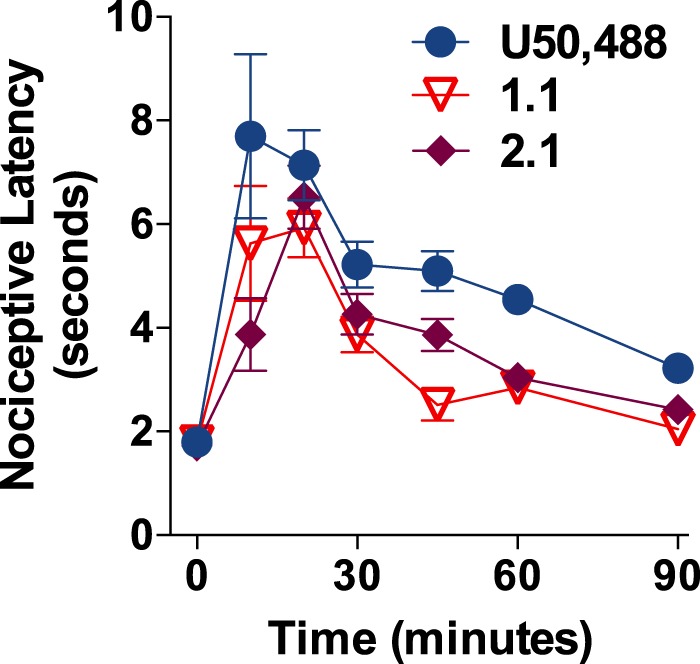
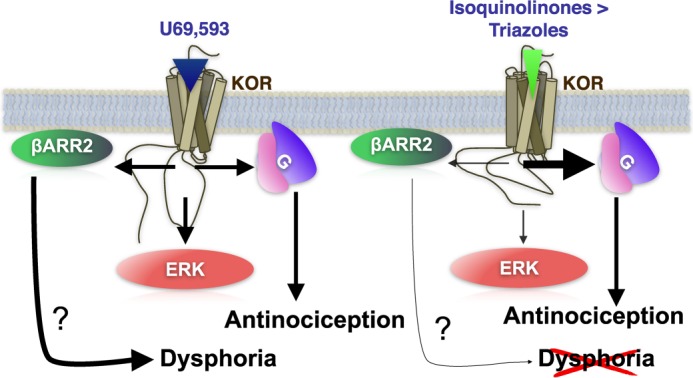
The kappa opioid receptor (KOR) is widely expressed in the CNS and can serve as a means to modulate pain perception, stress responses, and affective reward states. Therefore, the KOR has become a prominent drug discovery target toward treating pain, depression, and drug addiction. Agonists at KOR can promote G protein coupling and βarrestin2 recruitment as well as multiple downstream signaling pathways, including ERK1/2 MAPK activation. It has been suggested that the physiological effects of KOR activation result from different signaling cascades, with analgesia being G protein-mediated and dysphoria being mediated through βarrestin2 recruitment. Dysphoria associated with KOR activation limits the therapeutic potential in the use of KOR agonists as analgesics; therefore, it may be beneficial to develop KOR agonists that are biased toward G protein coupling and away from βarrestin2 recruitment. Here, we describe two classes of biased KOR agonists that potently activate G protein coupling but weakly recruit βarrestin2. These potent and functionally selective small molecule compounds may prove to be useful tools for refining the therapeutic potential of KOR-directed signaling in vivo.
Keywords: Arrestin; Brain; Drug Discovery; Dysphoria; ERK; G Protein-coupled Receptors (GPCR); Kappa Opioid Receptor; Opiate Opioid; Pain.
Figures

References
-
- Cox B. M. (2013) Recent developments in the study of opioid receptors. Mol. Pharmacol. 83, 723–728 - PubMed
-
- Chavkin C., James I. F., Goldstein A. (1982) Dynorphin is a specific endogenous ligand of the kappa opioid receptor. Science 215, 413–415 - PubMed
-
- Mansour A., Fox C. A., Burke S., Meng F., Thompson R. C., Akil H., Watson S. J. (1994) Mu, delta, and kappa opioid receptor mRNA expression in the rat CNS: an in situ hybridization study. J. Comp. Neurol. 350, 412–438 - PubMed
-
- Minami M., Satoh M. (1995) Molecular biology of the opioid receptors: structures, functions and distributions. Neurosci. Res. 23, 121–145 - PubMed
-
- Prather P. L., McGinn T. M., Claude P. A., Liu-Chen L. Y., Loh H. H., Law P. Y. (1995) Properties of a kappa-opioid receptor expressed in CHO cells: interaction with multiple G-proteins is not specific for any individual Gα subunit and is similar to that of other opioid receptors. Brain Res. Mol. Brain Res. 29, 336–346 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
