

Unique neuromyelitis optica pathology produced in naïve rats by intracerebral administration of NMO-IgG
- PMID: 24190619
- PMCID: PMC3954950
- DOI: 10.1007/s00401-013-1204-8
Unique neuromyelitis optica pathology produced in naïve rats by intracerebral administration of NMO-IgG
Abstract
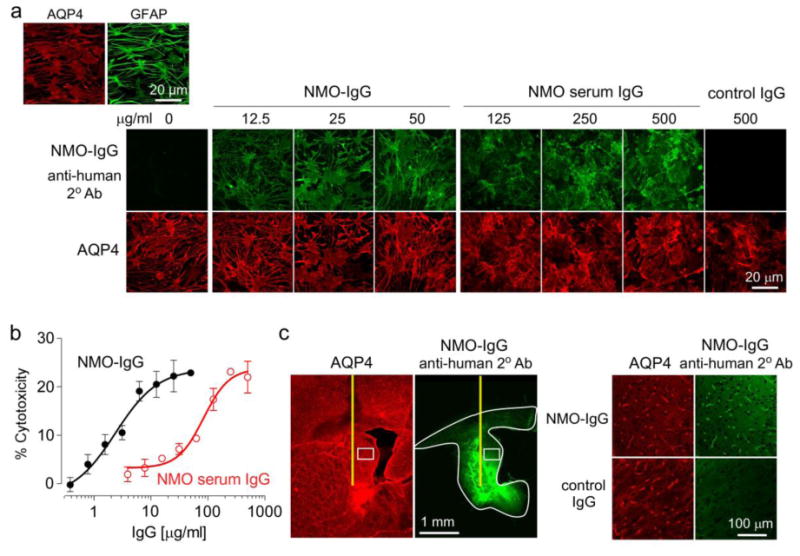
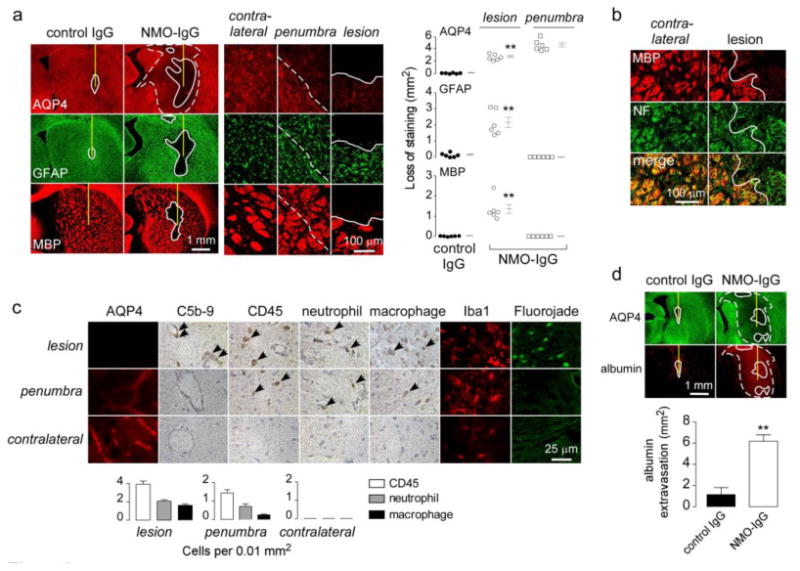
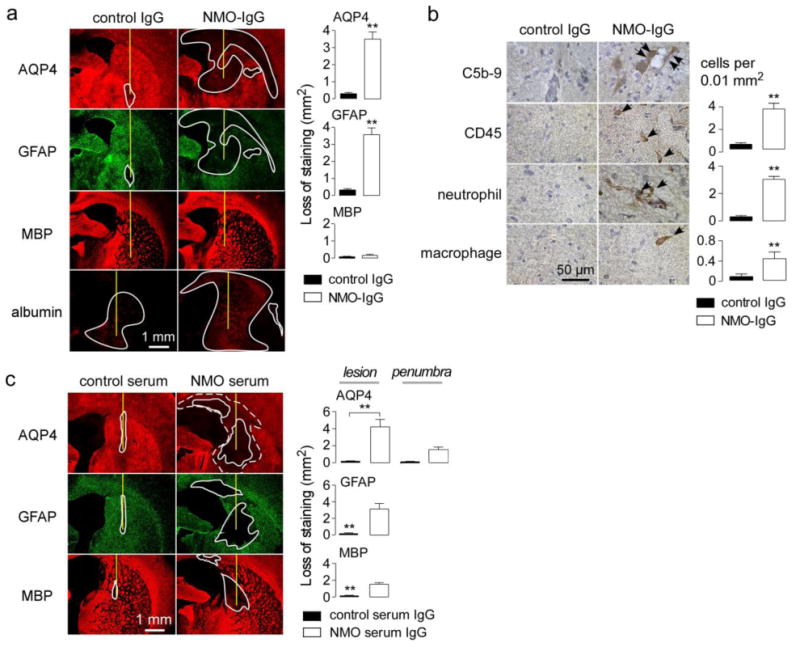
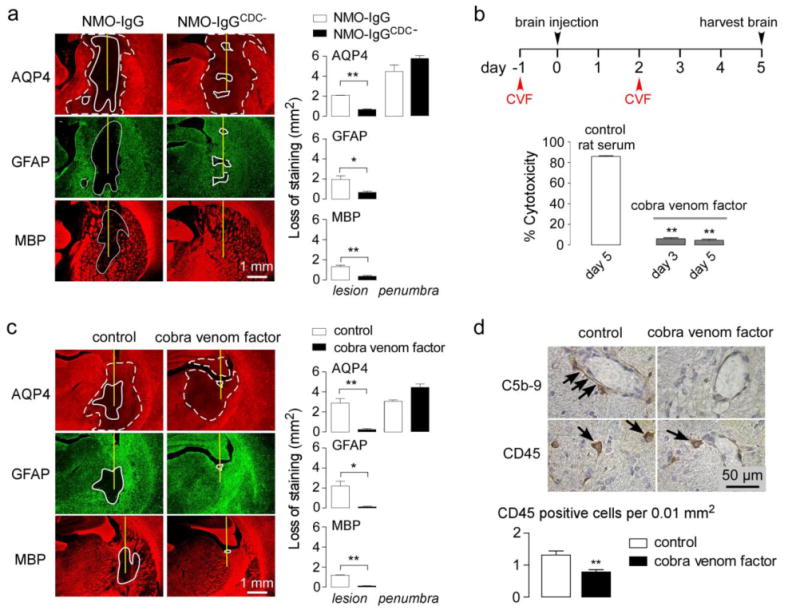
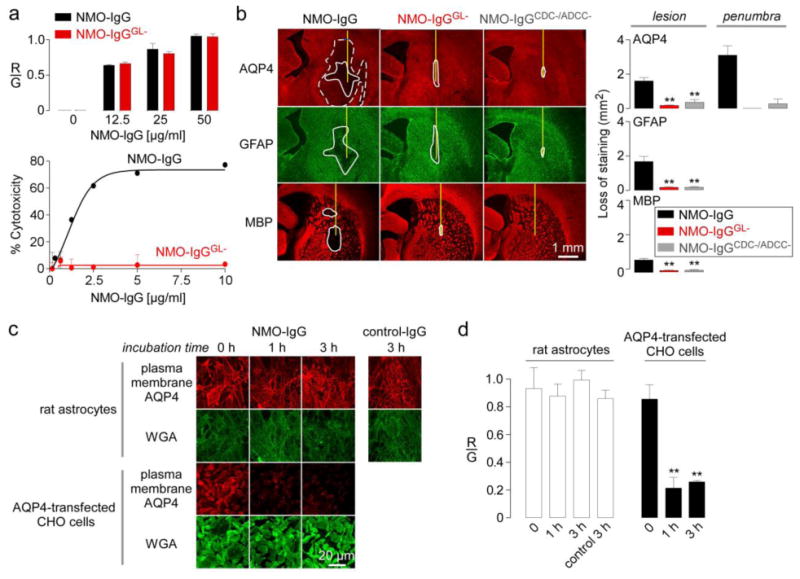
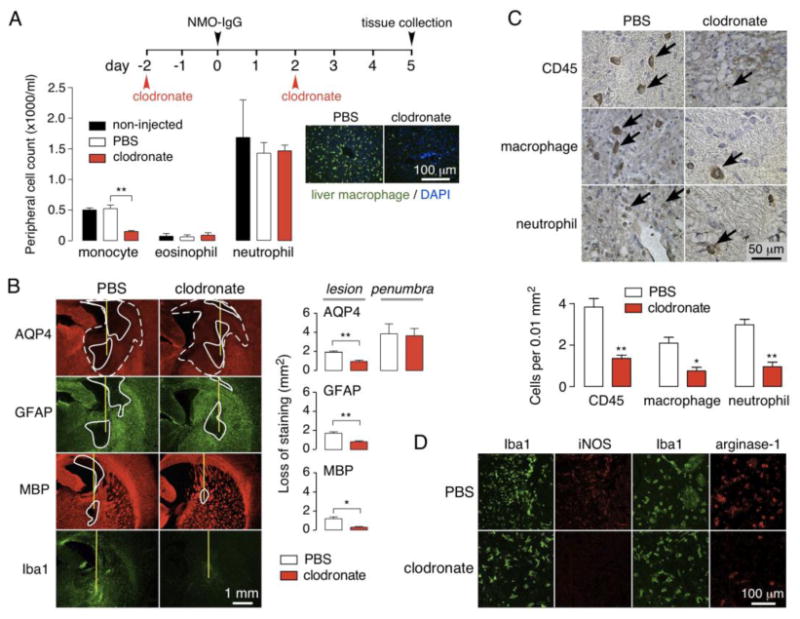
Animal models of neuromyelitis optica (NMO) are needed for elucidation of disease mechanisms and for testing new therapeutics. Prior animal models of NMO involved administration of human anti-aquaporin-4 immunoglobulin G antibody (NMO-IgG) to rats with pre-existing neuroinflammation, or to naïve mice supplemented with human complement. We report here the development of NMO pathology following passive transfer of NMO-IgG to naïve rats. A single intracerebral infusion of NMO-IgG to adult Lewis rats produced robust lesions around the needle track in 100 % of rats; at 5 days there was marked loss of aquaporin-4 (AQP4), glial fibrillary acidic protein (GFAP) and myelin, granulocyte and macrophage infiltration, vasculocentric complement deposition, blood-brain barrier disruption, microglial activation and neuron death. Remarkably, a distinct 'penumbra' was seen around lesions, with loss of AQP4 but not of GFAP or myelin. No lesions or penumbra were seen in rats receiving control IgG. The size of the main lesion with loss of myelin was greatly reduced in rats made complement-deficient by cobra venom factor or administered NMO-IgG lacking complement-dependent cytotoxicity (CDC) effector function. However, the penumbra was seen under these conditions, suggesting a complement-independent pathogenesis mechanism. The penumbra was absent with NMO-IgG lacking both CDC and antibody-dependent cellular cytotoxicity (ADCC) effector functions. Finally, lesion size was significantly reduced after macrophage depletion with clodronate liposomes. These results: (i) establish a robust, passive-transfer model of NMO in rats that does not require pre-existing neuroinflammation or complement administration; (ii) implicate ADCC as responsible for a unique type of pathology also seen in human NMO; and (iii) support a pathogenic role of macrophages in NMO.
Figures
References
-
- Bennett JL, Lam C, Kalluri SR, Saikali P, Bautista K, Dupree C, Glogowska M, Case D, Antel JP, Owens GP, Gilden D, Nessler S, Stadelmann C, Hemmer B. Intrathecal pathogenic anti-aquaporin-4 antibodies in early neuromyelitis optica. Ann Neurol. 2009;66(5):617–629. doi: 10.1002/ana.21802. - DOI - PMC - PubMed
-
- Bodega G, Suarez I, Lopez-Fernandez LA, Almonacid L, Zaballos A, Fernandez B. Possible implication of ciliary neurotrophic factor (CNTF) and beta-synuclein in the ammonia effect on cultured rat astroglial cells: a study using DNA and protein microarrays. Neurochem Int. 2006;48(8):729–738. doi:S0197-0186(06)00004-0. - PubMed
-
- Bush TG, Puvanachandra N, Horner CH, Polito A, Ostenfeld T, Svendsen CN, Mucke L, Johnson MH, Sofroniew MV. Leukocyte infiltration, neuronal degeneration, and neurite outgrowth after ablation of scar-forming, reactive astrocytes in adult transgenic mice. Neuron. 1999;23(2):297–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HL73856/HL/NHLBI NIH HHS/United States
- R01 EY013574/EY/NEI NIH HHS/United States
- DK86125/DK/NIDDK NIH HHS/United States
- R01 EB000415/EB/NIBIB NIH HHS/United States
- R01 DK035124/DK/NIDDK NIH HHS/United States
- DK72517/DK/NIDDK NIH HHS/United States
- EY13574/EY/NEI NIH HHS/United States
- DK35124/DK/NIDDK NIH HHS/United States
- R01 HL073856/HL/NHLBI NIH HHS/United States
- P30 DK072517/DK/NIDDK NIH HHS/United States
- RC1 DK086125/DK/NIDDK NIH HHS/United States
- EB00415/EB/NIBIB NIH HHS/United States
- R37 DK035124/DK/NIDDK NIH HHS/United States
- R37 EB000415/EB/NIBIB NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
