

The Aspergillus nidulans ATM kinase regulates mitochondrial function, glucose uptake and the carbon starvation response
- PMID: 24192833
- PMCID: PMC3887539
- DOI: 10.1534/g3.113.008607
The Aspergillus nidulans ATM kinase regulates mitochondrial function, glucose uptake and the carbon starvation response
Abstract
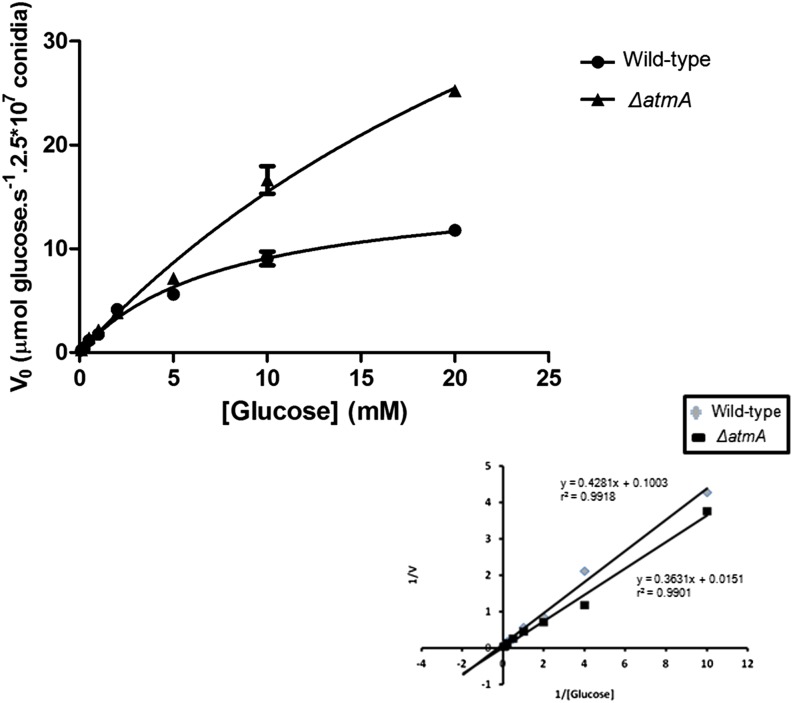
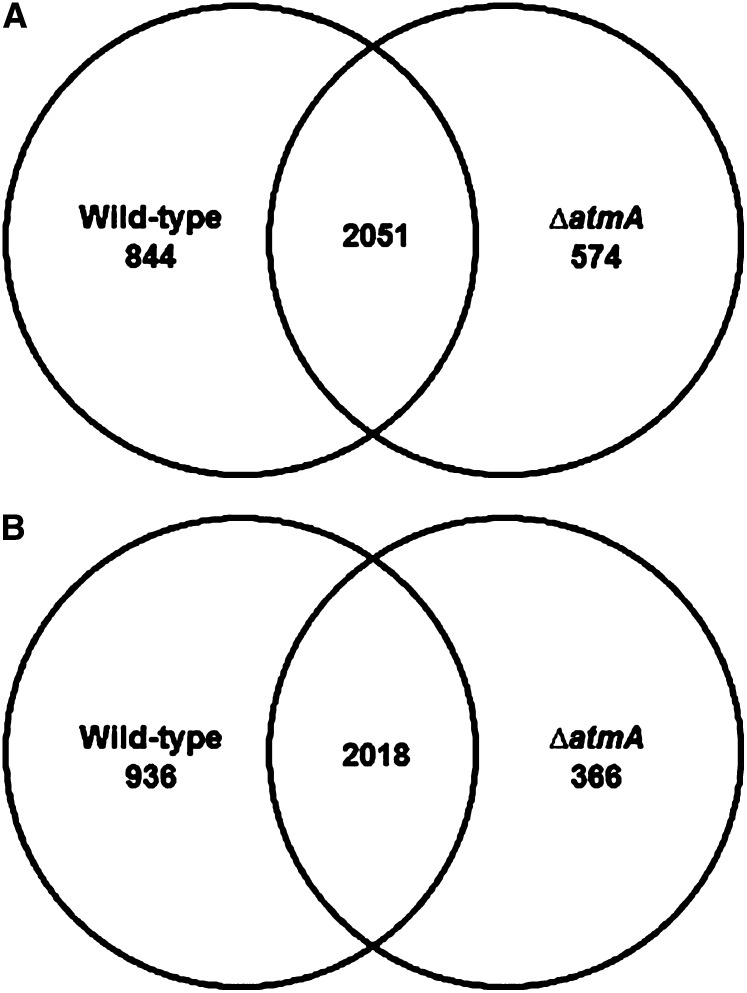
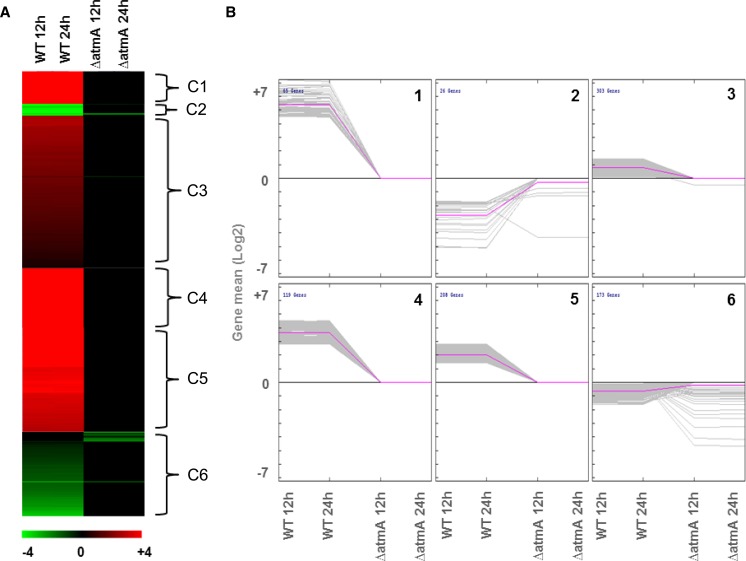
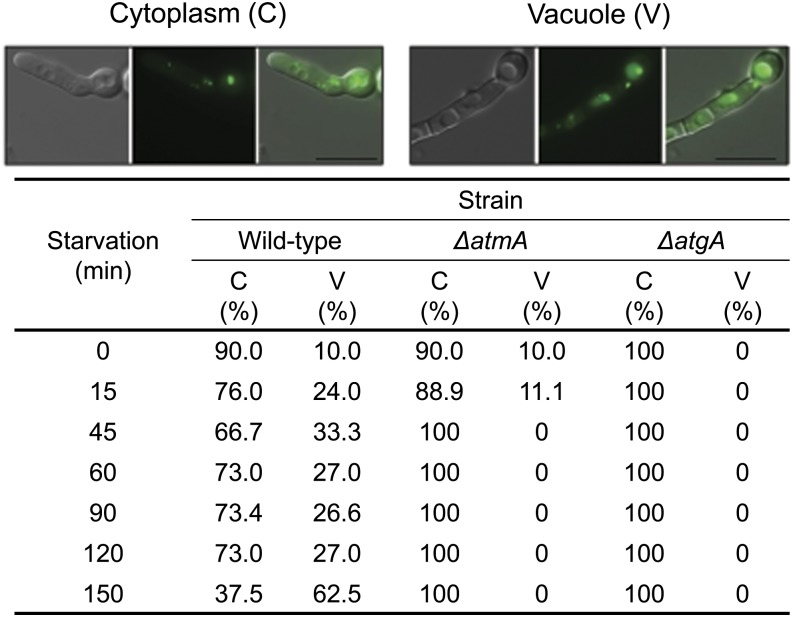
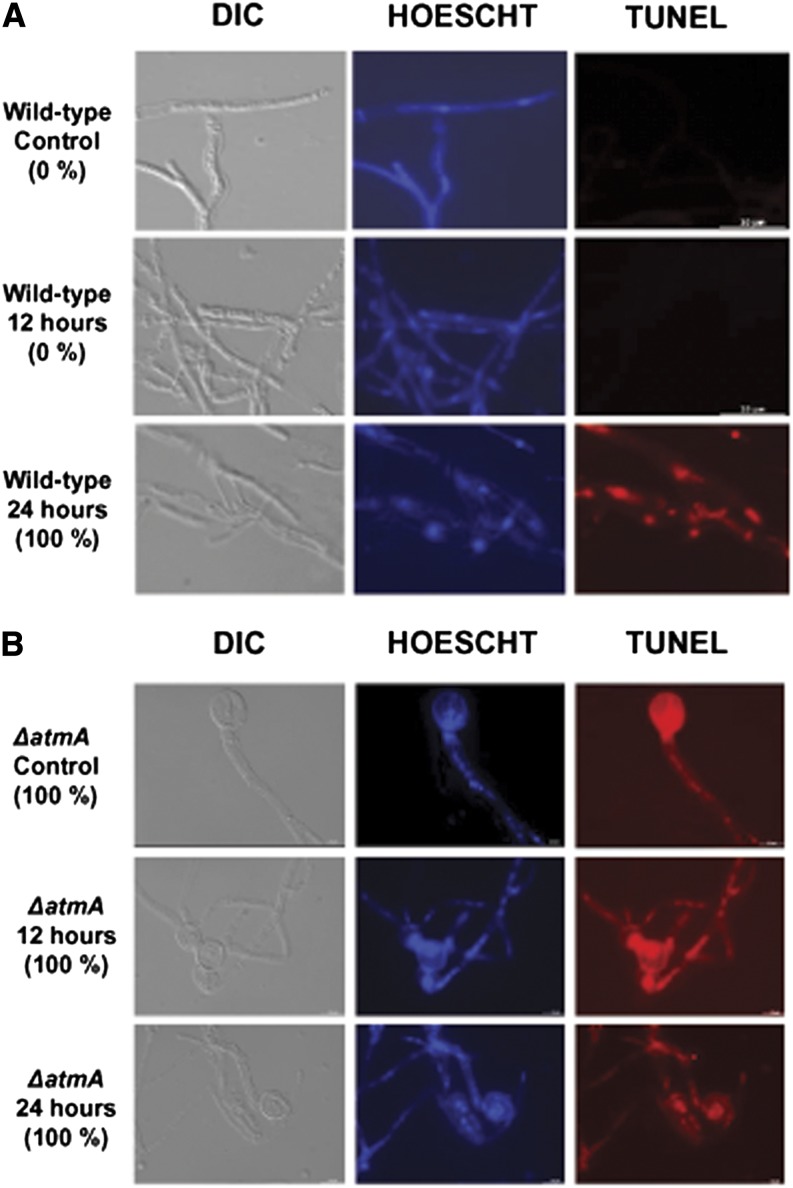
Mitochondria supply cellular energy and also perform a role in the adaptation to metabolic stress. In mammals, the ataxia-telangiectasia mutated (ATM) kinase acts as a redox sensor controlling mitochondrial function. Subsequently, transcriptomic and genetic studies were utilized to elucidate the role played by a fungal ATM homolog during carbon starvation. In Aspergillus nidulans, AtmA was shown to control mitochondrial function and glucose uptake. Carbon starvation responses that are regulated by target of rapamycin (TOR) were shown to be AtmA-dependent, including autophagy and hydrolytic enzyme secretion. AtmA also regulated a p53-like transcription factor, XprG, inhibiting starvation-induced XprG-dependent protease secretion and cell death. Thus, AtmA possibly represents a direct or indirect link between mitochondrial stress, metabolism, and growth through the influence of TOR and XprG function. The coordination of cell growth and division with nutrient availability is crucial for all microorganisms to successfully proliferate in a heterogeneous environment. Mitochondria supply cellular energy but also perform a role in the adaptation to metabolic stress and the cross-talk between prosurvival and prodeath pathways. The present study of Aspergillus nidulans demonstrated that AtmA also controlled mitochondrial mass, function, and oxidative phosphorylation, which directly or indirectly influenced glucose uptake. Carbon starvation responses, including autophagy, shifting metabolism to the glyoxylate cycle, and the secretion of carbon scavenging enzymes were AtmA-dependent. Transcriptomic profiling of the carbon starvation response demonstrated how TOR signaling and the retrograde response, which signals mitochondrial dysfunction, were directly or indirectly influenced by AtmA. The AtmA kinase was also shown to influence a p53-like transcription factor, inhibiting starvation-induced XprG-dependent protease secretion and cell death. Therefore, in response to metabolic stress, AtmA appears to perform a role in the regulation of TOR signaling, involving the retrograde and SnfA pathways. Thus, AtmA may represent a link between mitochondrial function and cell cycle or growth, possibly through the influence of the TOR and XprG function.
Keywords: ATM kinase; autophagy; cell death; glucose starvation.
Figures




References
-
- Banin S., Shieh S., Taya Y., Anderson C. W., Chessa L., et al. , 1998. Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science 281: 1674–1677. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous