

Release from meiotic arrest in ascidian eggs requires the activity of two phosphatases but not CaMKII
- PMID: 24194472
- PMCID: PMC3817944
- DOI: 10.1242/dev.096578
Release from meiotic arrest in ascidian eggs requires the activity of two phosphatases but not CaMKII
Abstract
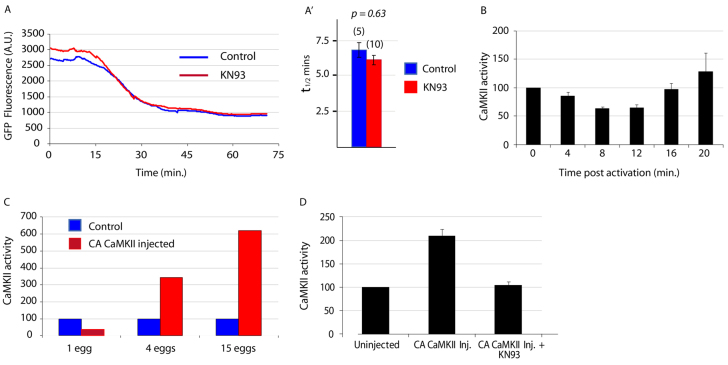
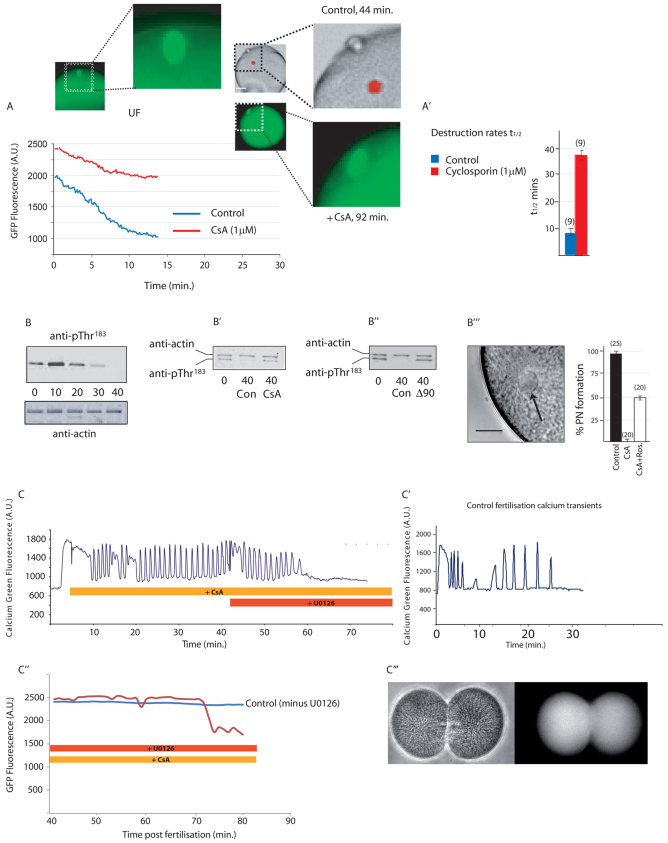
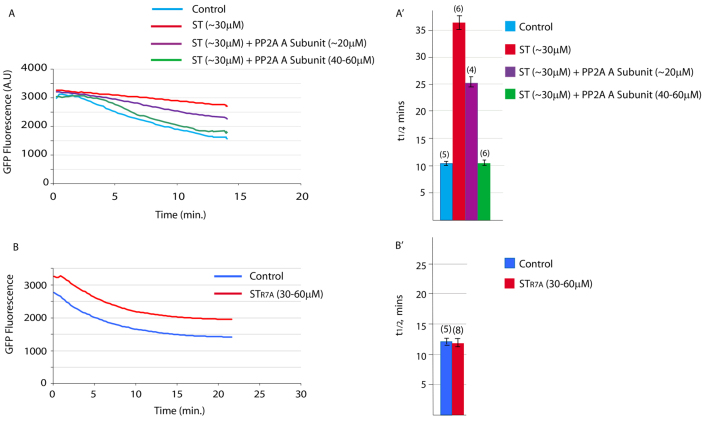
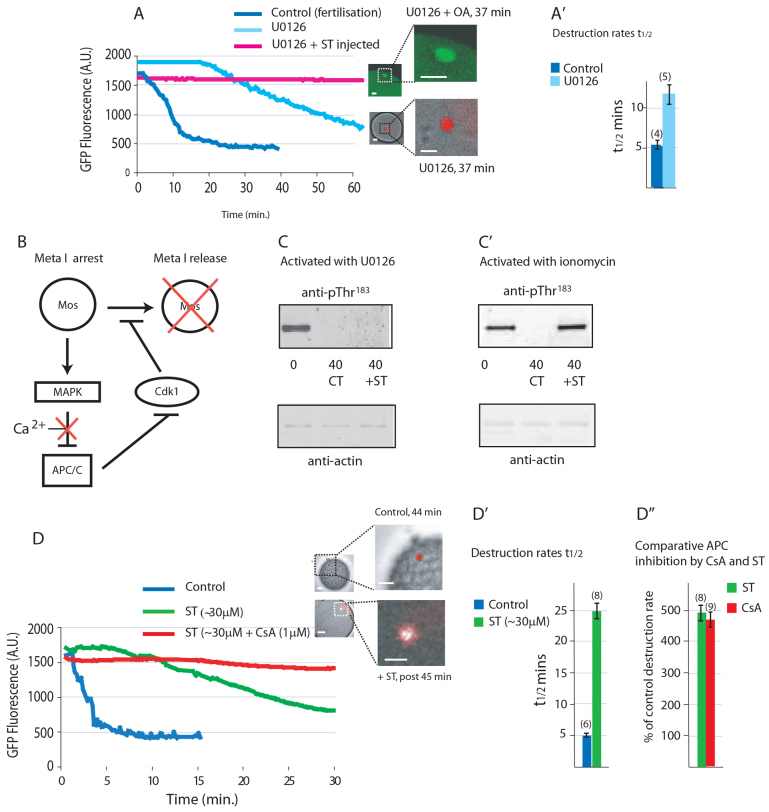
The fertilising sperm triggers a transient Ca(2+) increase that releases eggs from cell cycle arrest in the vast majority of animal eggs. In vertebrate eggs, Erp1, an APC/C(cdc20) inhibitor, links release from metaphase II arrest with the Ca(2+) transient and its degradation is triggered by the Ca(2+)-induced activation of CaMKII. By contrast, many invertebrate groups have mature eggs that arrest at metaphase I, and these species do not possess the CaMKII target Erp1 in their genomes. As a consequence, it is unknown exactly how cell cycle arrest at metaphase I is achieved and how the fertilisation Ca(2+) transient overcomes the arrest in the vast majority of animal species. Using live-cell imaging with a novel cyclin reporter to study cell cycle arrest and its release in urochordate ascidians, the closest living invertebrate group to the vertebrates, we have identified a new signalling pathway for cell cycle resumption in which CaMKII plays no part. Instead, we find that the Ca(2+)-activated phosphatase calcineurin (CN) is required for egg activation. Moreover, we demonstrate that parthenogenetic activation of metaphase I-arrested eggs by MEK inhibition, independent of a Ca(2+) increase, requires the activity of a second egg phosphatase: PP2A. Furthermore, PP2A activity, together with CN, is required for normal egg activation during fertilisation. As ascidians are a sister group of the vertebrates, we discuss these findings in relation to cell cycle arrest and egg activation in chordates.
Keywords: CaMKII; Calcineurin; Egg activation; Meiotic arrest; PP2A.
Figures

Similar articles
-
Calcium triggers exit from meiosis II by targeting the APC/C inhibitor XErp1 for degradation.Nature. 2005 Oct 13;437(7061):1048-52. doi: 10.1038/nature04093. Epub 2005 Aug 28. Nature. 2005. PMID: 16127448
-
Protein phosphatase 2A is essential to maintain meiotic arrest, and to prevent Ca2+ burst at spawning and eventual parthenogenesis in the larvacean Oikopleura dioica.Dev Biol. 2020 Apr 15;460(2):155-163. doi: 10.1016/j.ydbio.2019.12.005. Epub 2019 Dec 16. Dev Biol. 2020. PMID: 31857067
-
Calmodulin-dependent protein kinase II, and not protein kinase C, is sufficient for triggering cell-cycle resumption in mammalian eggs.J Cell Sci. 2005 Sep 1;118(Pt 17):3849-59. doi: 10.1242/jcs.02506. Epub 2005 Aug 9. J Cell Sci. 2005. PMID: 16091425
-
Cell cycle in ascidian eggs and embryos.Results Probl Cell Differ. 2011;53:153-69. doi: 10.1007/978-3-642-19065-0_8. Results Probl Cell Differ. 2011. PMID: 21630145 Review.
-
Mammalian egg activation: from Ca2+ spiking to cell cycle progression.Reproduction. 2005 Dec;130(6):813-23. doi: 10.1530/rep.1.00710. Reproduction. 2005. PMID: 16322541 Review.
Cited by
-
Aneuploidy in Oocytes Is Prevented by Sustained CDK1 Activity through Degron Masking in Cyclin B1.Dev Cell. 2019 Mar 11;48(5):672-684.e5. doi: 10.1016/j.devcel.2019.01.008. Epub 2019 Feb 7. Dev Cell. 2019. PMID: 30745144 Free PMC article.
-
Postovulatory cell death: why eggs die via apoptosis in biological species with external fertilization.J Reprod Dev. 2018 Feb 27;64(1):1-6. doi: 10.1262/jrd.2017-100. Epub 2017 Oct 27. J Reprod Dev. 2018. PMID: 29081453 Free PMC article. Review.
-
Inhibition of calcineurin by FK506 stimulates germinal vesicle breakdown of mouse oocytes in hypoxanthine-supplemented medium.PeerJ. 2017 Feb 23;5:e3032. doi: 10.7717/peerj.3032. eCollection 2017. PeerJ. 2017. PMID: 28243539 Free PMC article.
-
Gamete activation: basic knowledge and clinical applications.Hum Reprod Update. 2016 Jun;22(4):420-39. doi: 10.1093/humupd/dmw014. Epub 2016 Jun 8. Hum Reprod Update. 2016. PMID: 27278231 Free PMC article. Review.
-
Signaling Enzymes Required for Sperm Maturation and Fertilization in Mammals.Front Cell Dev Biol. 2019 Dec 18;7:341. doi: 10.3389/fcell.2019.00341. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31921853 Free PMC article. Review.
References
-
- Amiel A., Leclère L., Robert L., Chevalier S., Houliston E. (2009). Conserved functions for Mos in eumetazoan oocyte maturation revealed by studies in a cnidarian. Curr. Biol. 19, 305–311 - PubMed
-
- Chang H. Y., Minahan K., Merriman J. A., Jones K. T. (2009). Calmodulin-dependent protein kinase gamma 3 (CamKIIgamma3) mediates the cell cycle resumption of metaphase II eggs in mouse. Development 136, 4077–4081 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
