

pH of drinking water influences the composition of gut microbiome and type 1 diabetes incidence
- PMID: 24194504
- PMCID: PMC3900548
- DOI: 10.2337/db13-0981
pH of drinking water influences the composition of gut microbiome and type 1 diabetes incidence
Abstract
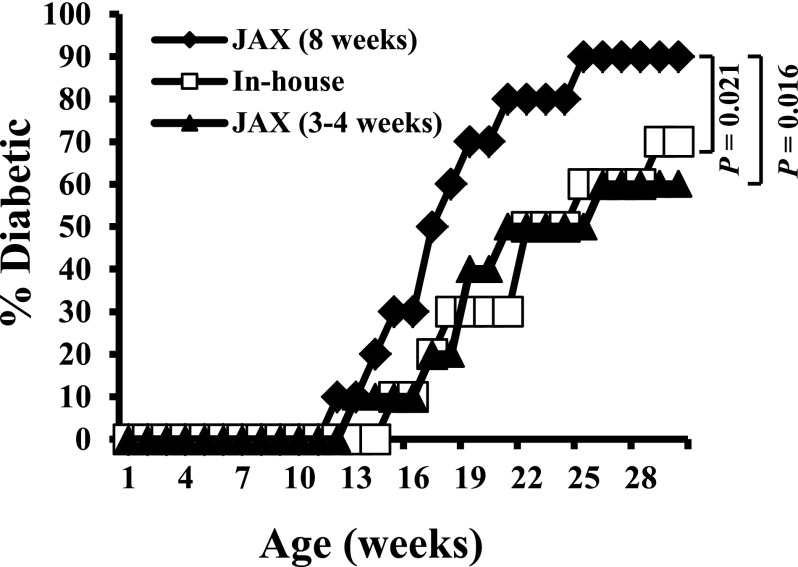
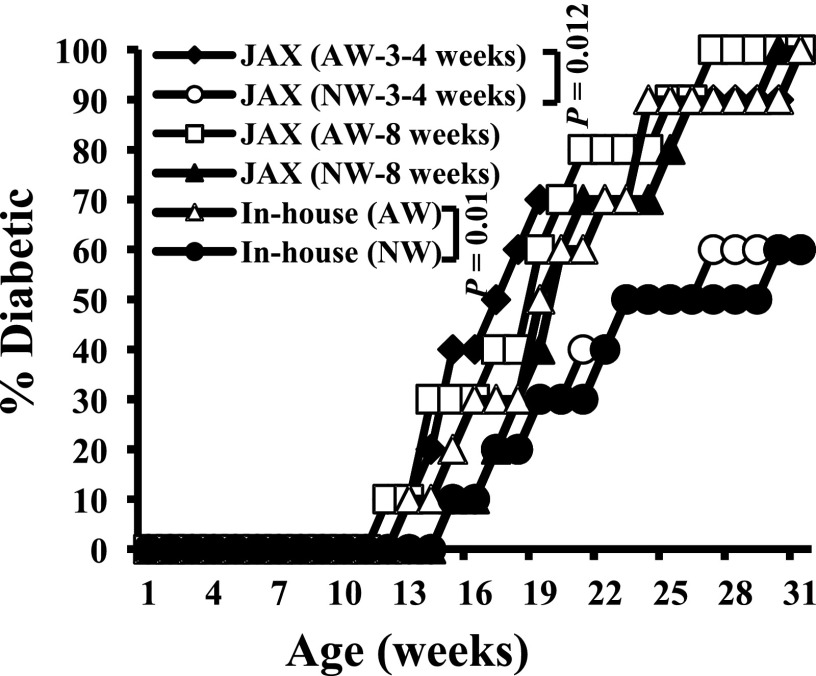
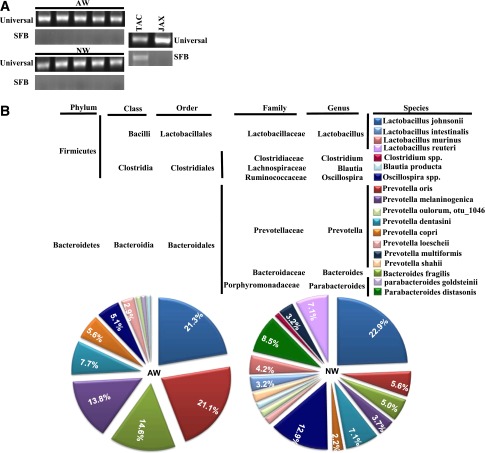
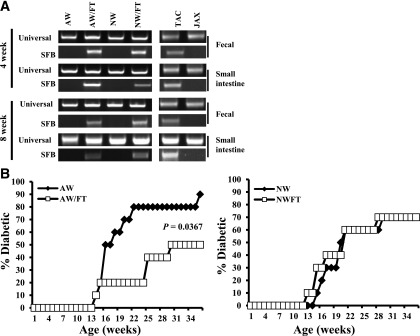
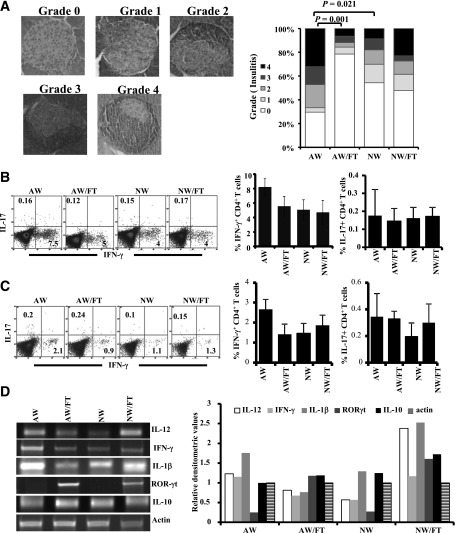
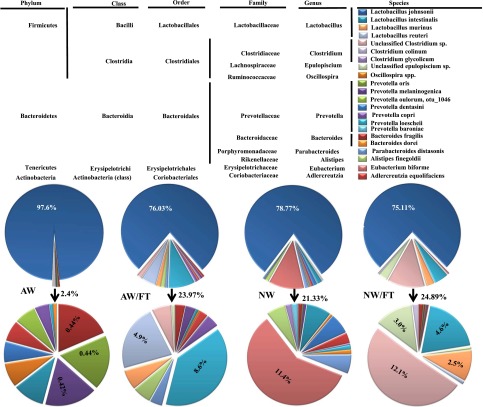
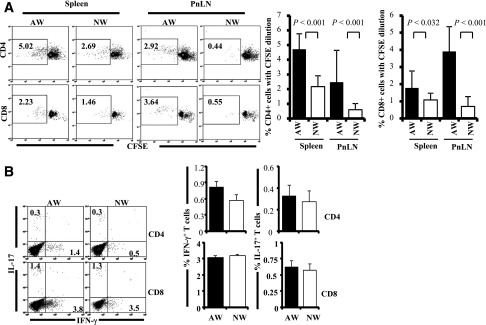
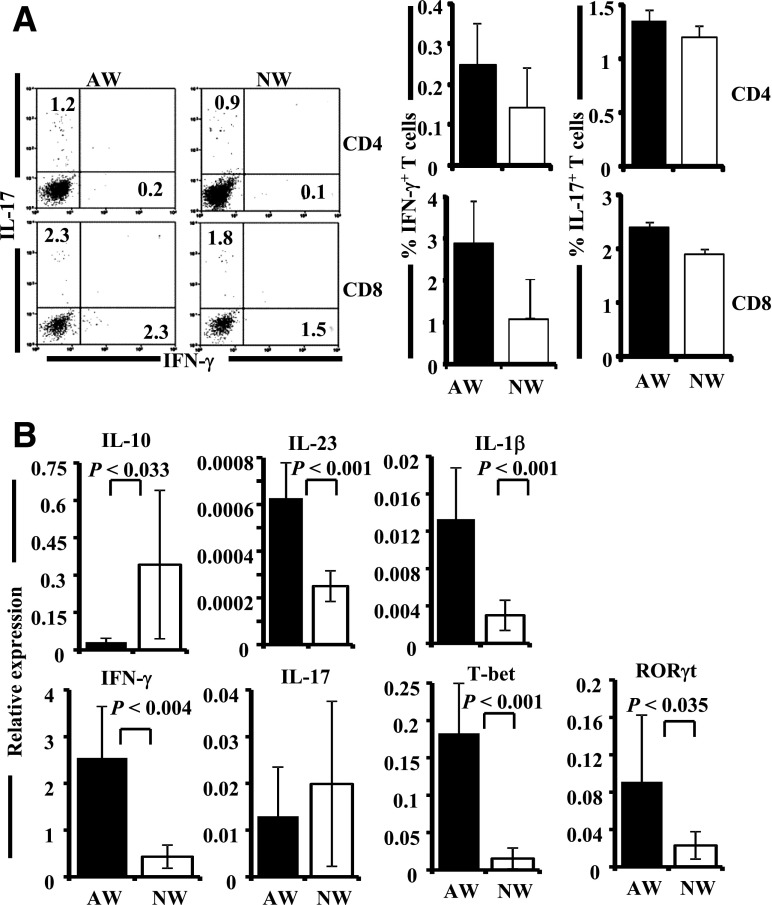
Nonobese diabetic (NOD) mice spontaneously develop type 1 diabetes (T1D), progression of which is similar to that in humans, and therefore are widely used as a model for understanding the immunological basis of this disease. The incidence of T1D in NOD mice is influenced by the degree of cleanliness of the mouse colony and the gut microflora. In this report, we show that the T1D incidence and rate of disease progression are profoundly influenced by the pH of drinking water, which also affects the composition and diversity of commensal bacteria in the gut. Female NOD mice that were maintained on acidic pH water (AW) developed insulitis and hyperglycemia rapidly compared with those on neutral pH water (NW). Interestingly, forced dysbiosis by segmented filamentous bacteria (SFB)-positive fecal transfer significantly suppressed the insulitis and T1D incidence in mice that were on AW but not in those on NW. The 16S rDNA-targeted pyrosequencing revealed a significant change in the composition and diversity of gut flora when the pH of drinking water was altered. Importantly, autoantigen-specific T-cell frequencies in the periphery and proinflammatory cytokine response in the intestinal mucosa are significantly higher in AW-recipient mice compared with their NW counterparts. These observations suggest that pH of drinking water affects the composition of gut microflora, leading to an altered autoimmune response and T1D incidence in NOD mice.
Figures
Comment in
-
Comment on Sofi et al. pH of Drinking Water Influences the Composition of Gut Microbiome and Type 1 Diabetes Incidence. Diabetes 2014;63:632-644.Diabetes. 2015 Aug;64(8):e19. doi: 10.2337/db15-0321. Diabetes. 2015. PMID: 26207042 Free PMC article. No abstract available.
-
Response to Comment on Sofi et al. pH of Drinking Water Influences the Composition of Gut Microbiome and Type 1 Diabetes Incidence. Diabetes 2014;63:632-644.Diabetes. 2015 Aug;64(8):e20-1. doi: 10.2337/db15-0554. Diabetes. 2015. PMID: 26207043 No abstract available.
References
-
- Rook GA. Hygiene hypothesis and autoimmune diseases. Clin Rev Allergy Immunol 2012;42:5–15 - PubMed
-
- Bach JF. Protective role of infections and vaccinations on autoimmune diseases. J Autoimmun 2001;16:347–353 - PubMed
-
- Pozzilli P, Signore A, Williams AJ, Beales PE. NOD mouse colonies around the world—recent facts and figures. Immunol Today 1993;14:193–196 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
