

Regulation of drought tolerance by the F-box protein MAX2 in Arabidopsis
- PMID: 24198318
- PMCID: PMC3875819
- DOI: 10.1104/pp.113.226837
Regulation of drought tolerance by the F-box protein MAX2 in Arabidopsis
Abstract
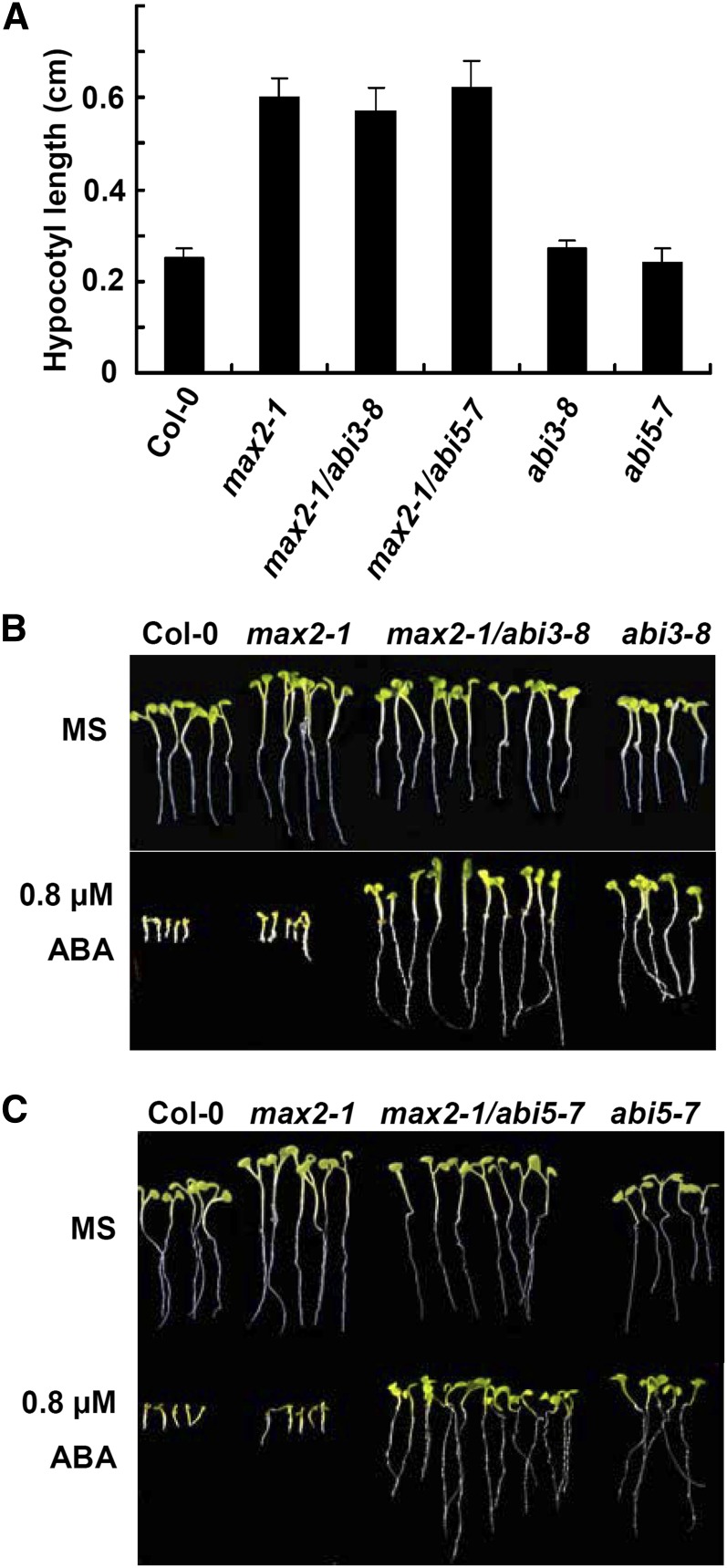
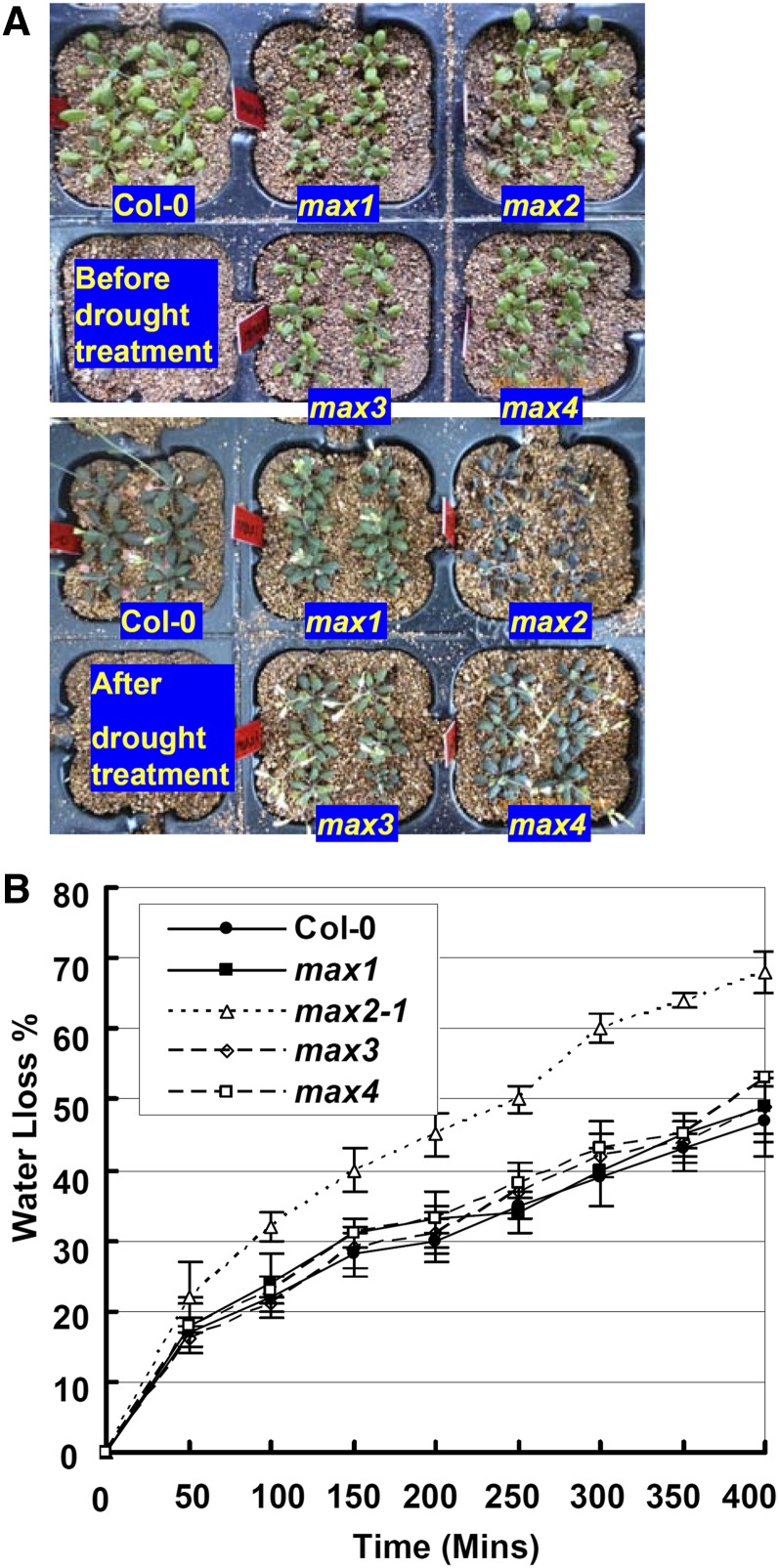
MAX2 (for MORE AXILLARY GROWTH2) has been shown to regulate diverse biological processes, including plant architecture, photomorphogenesis, senescence, and karrikin signaling. Although karrikin is a smoke-derived abiotic signal, a role for MAX2 in abiotic stress response pathways is least investigated. Here, we show that the max2 mutant is strongly hypersensitive to drought stress compared with wild-type Arabidopsis (Arabidopsis thaliana). Stomatal closure of max2 was less sensitive to abscisic acid (ABA) than that of the wild type. Cuticle thickness of max2 was significantly thinner than that of the wild type. Both of these phenotypes of max2 mutant plants correlate with the increased water loss and drought-sensitive phenotype. Quantitative real-time reverse transcription-polymerase chain reaction analyses showed that the expression of stress-responsive genes and ABA biosynthesis, catabolism, transport, and signaling genes was impaired in max2 compared with wild-type seedlings in response to drought stress. Double mutant analysis of max2 with the ABA-insensitive mutants abi3 and abi5 indicated that MAX2 may function upstream of these genes. The expression of ABA-regulated genes was enhanced in imbibed max2 seeds. In addition, max2 mutant seedlings were hypersensitive to ABA and osmotic stress, including NaCl, mannitol, and glucose. Interestingly, ABA, osmotic stress, and drought-sensitive phenotypes were restricted to max2, and the strigolactone biosynthetic pathway mutants max1, max3, and max4 did not display any defects in these responses. Taken together, these results uncover an important role for MAX2 in plant responses to abiotic stress conditions.
Figures









Similar articles
-
The Arabidopsis F-box protein FOF2 regulates ABA-mediated seed germination and drought tolerance.Plant Sci. 2020 Dec;301:110643. doi: 10.1016/j.plantsci.2020.110643. Epub 2020 Aug 28. Plant Sci. 2020. PMID: 33218620
-
MAX2 affects multiple hormones to promote photomorphogenesis.Mol Plant. 2012 May;5(3):750-62. doi: 10.1093/mp/sss029. Epub 2012 Mar 30. Mol Plant. 2012. PMID: 22466576
-
The FBA motif-containing protein AFBA1 acts as a novel positive regulator of ABA response in Arabidopsis.Plant Cell Physiol. 2017 Mar 1;58(3):574-586. doi: 10.1093/pcp/pcx003. Plant Cell Physiol. 2017. PMID: 28184867
-
Integration of light and ABA signaling pathways to combat drought stress in plants.Plant Cell Rep. 2023 May;42(5):829-841. doi: 10.1007/s00299-023-02999-7. Epub 2023 Mar 12. Plant Cell Rep. 2023. PMID: 36906730 Review.
-
Hormonal regulation of plant primary metabolism under drought.J Exp Bot. 2024 Mar 14;75(6):1714-1725. doi: 10.1093/jxb/erad358. J Exp Bot. 2024. PMID: 37712613 Review.
Cited by
-
SCF E3 ligase PP2-B11 plays a positive role in response to salt stress in Arabidopsis.J Exp Bot. 2015 Aug;66(15):4683-97. doi: 10.1093/jxb/erv245. Epub 2015 Jun 2. J Exp Bot. 2015. PMID: 26041321 Free PMC article.
-
Catabolism of strigolactones by a carboxylesterase.Nat Plants. 2021 Nov;7(11):1495-1504. doi: 10.1038/s41477-021-01011-y. Epub 2021 Nov 11. Nat Plants. 2021. PMID: 34764442
-
Strigolactones enhance root-knot nematode (Meloidogyne graminicola) infection in rice by antagonizing the jasmonate pathway.New Phytol. 2019 Oct;224(1):454-465. doi: 10.1111/nph.15953. Epub 2019 Jul 2. New Phytol. 2019. PMID: 31125438 Free PMC article.
-
Control of ABA Signaling and Crosstalk with Other Hormones by the Selective Degradation of Pathway Components.Int J Mol Sci. 2021 Apr 28;22(9):4638. doi: 10.3390/ijms22094638. Int J Mol Sci. 2021. PMID: 33924944 Free PMC article. Review.
-
SUPPRESSOR OF MAX2 LIKE 6, 7, and 8 Interact with DDB1 BINDING WD REPEAT DOMAIN HYPERSENSITIVE TO ABA DEFICIENT 1 to Regulate the Drought Tolerance and Target SUCROSE NONFERMENTING 1 RELATED PROTEIN KINASE 2.3 to Abscisic Acid Response in Arabidopsis.Biomolecules. 2023 Sep 18;13(9):1406. doi: 10.3390/biom13091406. Biomolecules. 2023. PMID: 37759806 Free PMC article.
References
-
- Booker J, Auldridge M, Wills S, McCarty D, Klee H, Leyser O. (2004) MAX3/CCD7 is a carotenoid cleavage dioxygenase required for the synthesis of a novel plant signaling molecule. Curr Biol 14: 1232–1238 - PubMed
-
- Booker J, Sieberer T, Wright W, Williamson L, Willett B, Stirnberg P, Turnbull C, Srinivasan M, Goddard P, Leyser O. (2005) MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Dev Cell 8: 443–449 - PubMed
-
- Brandt B, Brodsky DE, Xue S, Negi J, Iba K, Kangasjärvi J, Ghassemian M, Stephan AB, Hu H, Schroeder JI. (2012) Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proc Natl Acad Sci USA 109: 10593–10598 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
