

A local source of FGF initiates development of the unmyelinated lineage of sensory neurons
- PMID: 24198358
- PMCID: PMC6618424
- DOI: 10.1523/JNEUROSCI.1090-13.2013
A local source of FGF initiates development of the unmyelinated lineage of sensory neurons
Abstract
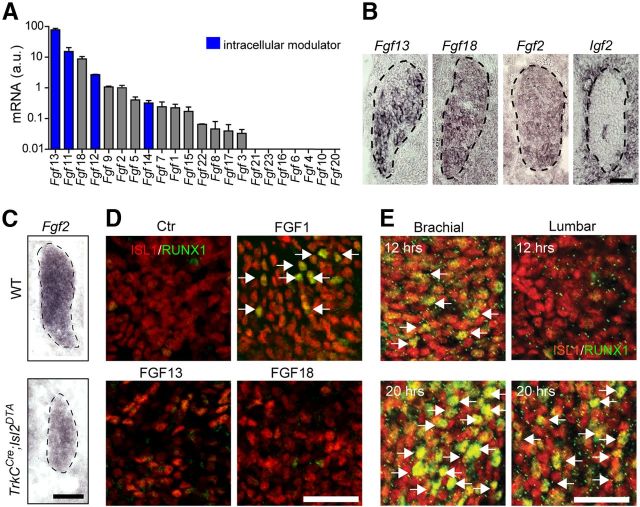
The principle by which unmyelinated primary sensory neurons transducing thermal, itch and pain perception are specified in early development is unknown. These classes of sensory neurons diversify from a common population of late-born neurons, which initiate expression of Runt homology domain transcription factor RUNX1 and the nerve growth factor receptor TrkA. Here, we report that signals emanating from within the mouse dorsal root ganglion mediated partly by early-born neurons destined to a myelinated phenotype participate in fating late-born RUNX1(+)/TrkA(+) neurons. Inductive factors include FGFs via activation of FGF receptor 1 (FGFR1). Consistently, FGF2 is sufficient to induce expression of RUNX1, and Fgfr1 conditional mutant mice display deficits in fating of the common population of late-born RUNX1(+)/TrkA(+) neurons that develop into unmyelinated neurons. Thus, the distinct lineages of sensory neurons are acquired in response to increasing FGF levels provided by a rising number of born neurons.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous