

Novel mutations in Lrp6 orthologs in mouse and human neural tube defects affect a highly dosage-sensitive Wnt non-canonical planar cell polarity pathway
- PMID: 24203697
- PMCID: PMC3943515
- DOI: 10.1093/hmg/ddt558
Novel mutations in Lrp6 orthologs in mouse and human neural tube defects affect a highly dosage-sensitive Wnt non-canonical planar cell polarity pathway
Erratum in
- Hum Mol Genet. 2014 Aug 1;23(15):4185
Abstract
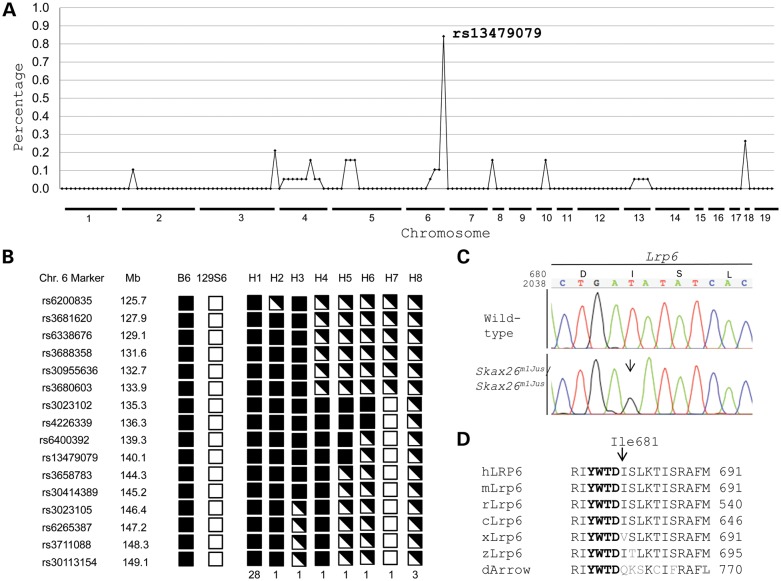
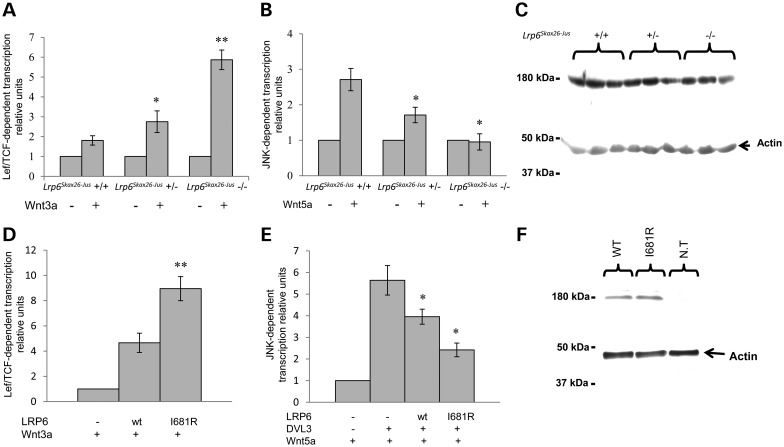
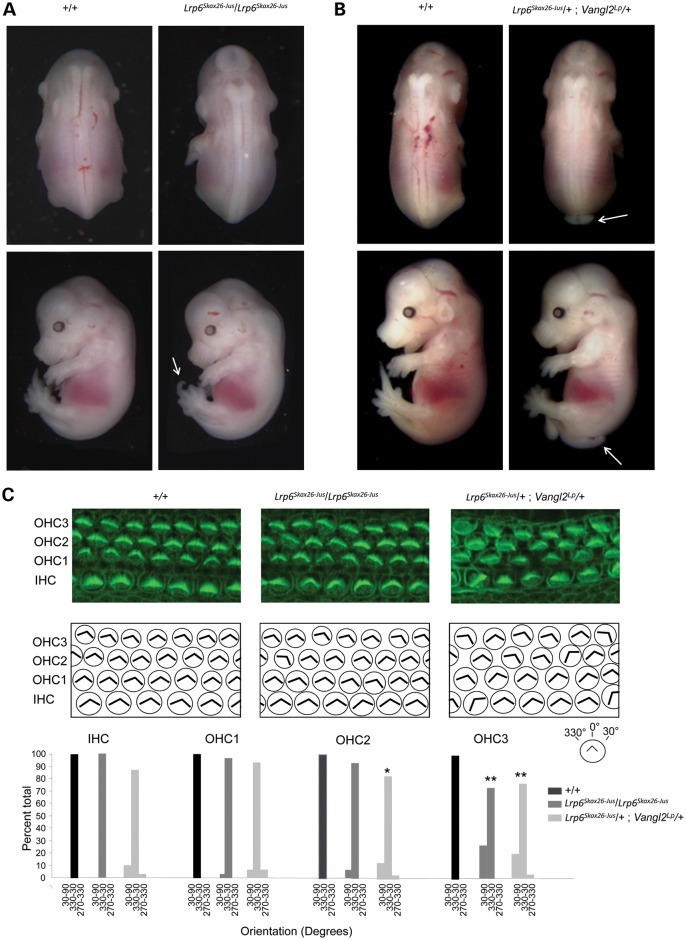
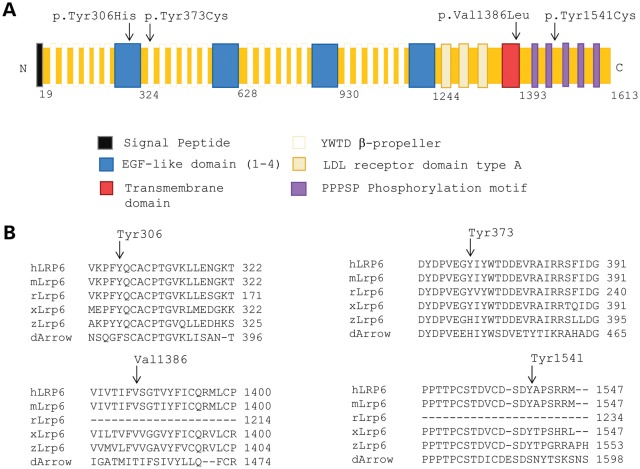
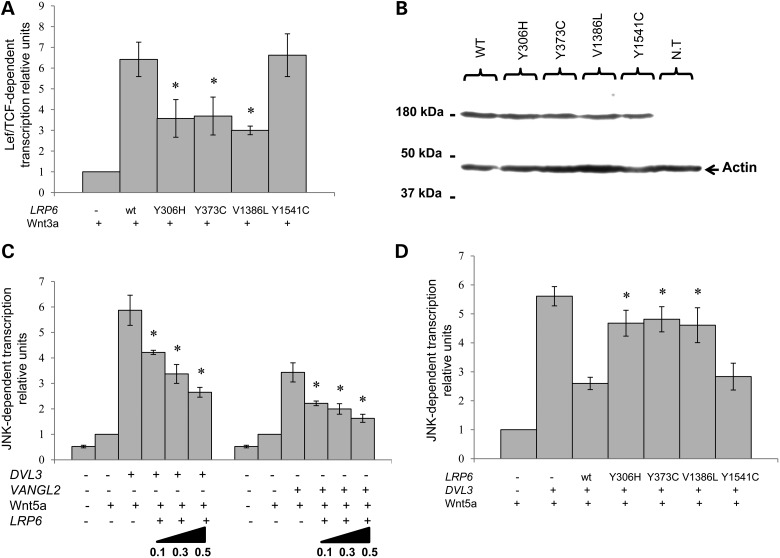
Wnt signaling has been classified as canonical Wnt/β-catenin-dependent or non-canonical planar cell polarity (PCP) pathway. Misregulation of either pathway is linked mainly to cancer or neural tube defects (NTDs), respectively. Both pathways seem to antagonize each other, and recent studies have implicated a number of molecular switches that activate one pathway while simultaneously inhibiting the other thereby partially mediating this antagonism. The lipoprotein receptor-related protein Lrp6 is crucial for the activation of the Wnt/β-catenin pathway, but its function in Wnt/PCP signaling remains largely unknown. In this study, we investigate the role of Lrp6 as a molecular switch between both Wnt pathways in a novel ENU mouse mutant of Lrp6 (Skax26(m1Jus)) and in human NTDs. We demonstrate that Skax26(m1Jus) represents a hypermorphic allele of Lrp6 with increased Wnt canonical and abolished PCP-induced JNK activities. We also show that Lrp6(Skax26-Jus) genetically interacts with a PCP mutant (Vangl2(Lp)) where double heterozygotes showed an increased frequency of NTDs and defects in cochlear hair cells' polarity. Importantly, our study also demonstrates the association of rare and novel missense mutations in LRP6 that is an inhibitor rather than an activator of the PCP pathway with human NTDs. We show that three LRP6 mutations in NTDs led to a reduced Wnt canonical activity and enhanced PCP signaling. Our data confirm an inhibitory role of Lrp6 in PCP signaling in neurulation and indicate the importance of a tightly regulated and highly dosage-sensitive antagonism between both Wnt pathways in this process.
Figures
References
-
- Niehrs C. The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol. 2012;13:767–779. - PubMed
-
- Montcouquiol M., Crenshaw E.B., 3rd, Kelley M.W. Noncanonical Wnt signaling and neural polarity. Annu. Rev. Neurosci. 2006;29:363–386. - PubMed
-
- Tada M., Heisenberg C.-P. Convergent extension: using collective cell migration and cell intercalation to shape embryos. Dev. Camb. Engl. 2012;139:3897–3904. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
